










組學(xué)數(shù)據(jù)向來是高質(zhì)量期刊較為看重的假說驗(yàn)證證據(jù),涉及的多組學(xué)實(shí)驗(yàn)環(huán)節(jié)越豐富,對于現(xiàn)象機(jī)制的探究往往更為完整和具體。在植物抗病機(jī)制研究領(lǐng)域,測序和代謝組分析手段已非罕見,要尋找新的創(chuàng)新點(diǎn)和研究亮點(diǎn),還需要再挖掘其他角度。
本期優(yōu)秀客戶文章解讀,看磷酸化蛋白質(zhì)組和納米材料兩大熱點(diǎn),如何為這篇發(fā)表于Journal of Cleaner Production的植物方向課題加分!

煙草青枯病由土傳病原菌R. solanacearum侵染煙草植物引起,是煙草種植中一種嚴(yán)重的病害,對煙草的產(chǎn)量和質(zhì)量造成了顯著影響。R. solanacearum通過分泌胞外多糖I導(dǎo)致木質(zhì)部物理堵塞,最終導(dǎo)致感病植物枯萎。
傳統(tǒng)煙草青枯病控制方法以化學(xué)藥劑根部灌溉為主,環(huán)保和控制效率不足。近年來,納米材料因其獨(dú)特的物理化學(xué)性質(zhì)和在農(nóng)業(yè)領(lǐng)域的潛在應(yīng)用價(jià)值,逐漸成為研究熱點(diǎn)。本研究選擇具有良好抗菌性能的鉍(Bi)和能增強(qiáng)植物抗逆性硒(Se)兩種元素作為切入點(diǎn),借助納米材料可穿過生物屏障并在施用于根部或葉片后擴(kuò)散到植物維管系統(tǒng)內(nèi)的能力,研究設(shè)計(jì)并合成了一種新型的鉍-硒納米復(fù)合簇(Bi-Se NPs),旨在探索其對煙草青枯病的防治效果及其作用機(jī)制。

研究對象:R. solanacearum菌株、煙草植株、鉍-硒納米復(fù)合簇
技術(shù)方法:非靶代謝組、真核轉(zhuǎn)錄組、RT-qPCR、磷酸化修飾蛋白質(zhì)組等
研究思路:
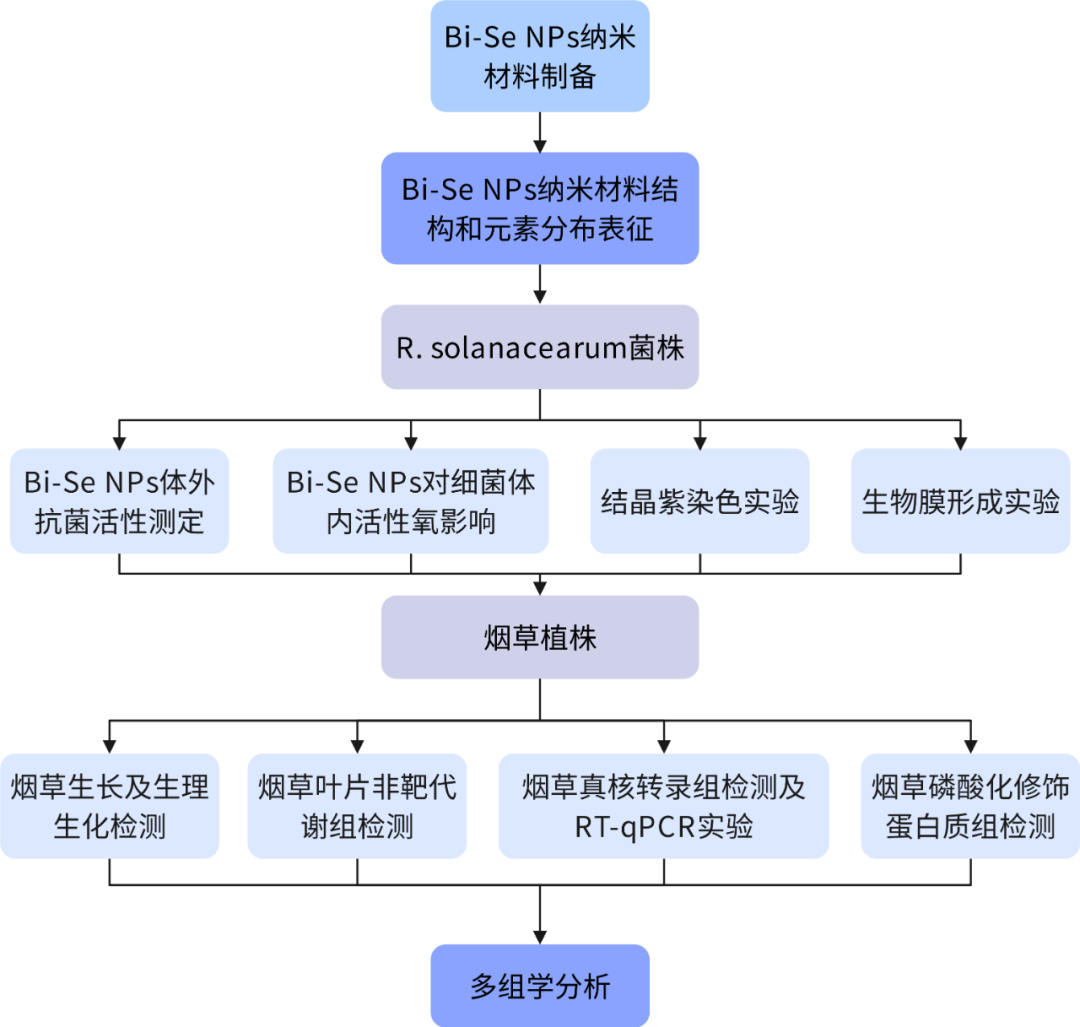
技術(shù)路線圖
1.Bi-Se NPs納米復(fù)合簇表征
在氮?dú)獗Wo(hù)下,Bi NPs和Se NPs通過靜電相互作用結(jié)合形成Bi-Se NPs。透射電子顯微鏡(TEM)和掃描電子顯微鏡(SEM)觀察結(jié)果顯示Bi NPs呈片狀結(jié)構(gòu),Se NPs呈均勻球形吸附在Bi NPs結(jié)構(gòu)表面且分布均勻(圖1A-C)。XPS和XRD均顯示出鉍和硒的特征峰(圖1G-J)。
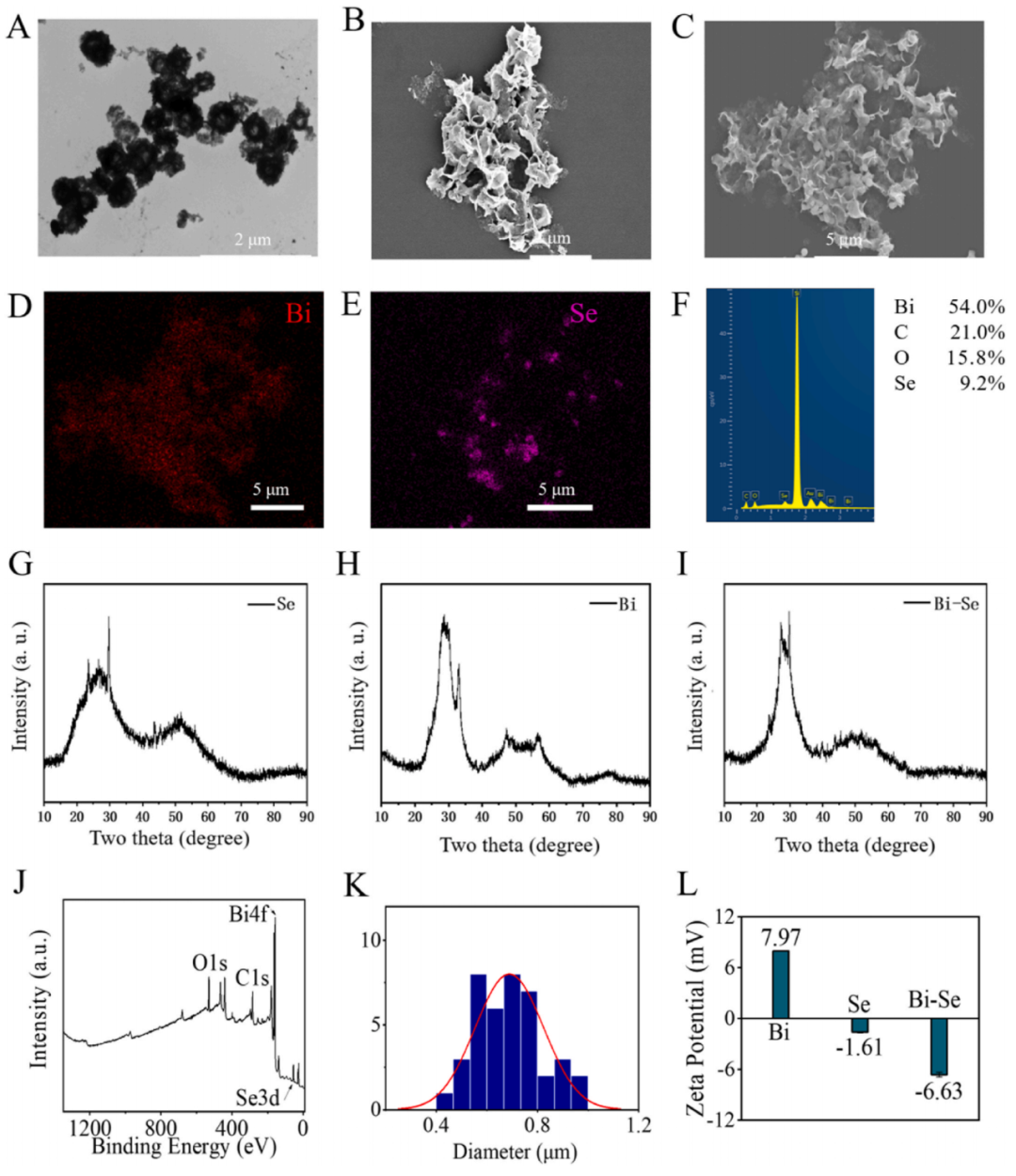
圖1. Bi-Se NPs的制備與表征
2.Bi-Se NPs的體外抗菌特性
Bi NPs或Se NPs單獨(dú)處理對R. solanacearum抗菌活性不明顯,相比之下,Bi-Se NPs有效抑制了細(xì)菌的增殖,菌落總數(shù)減少,并顯著提高了抗菌效果(圖2B)。此外,采用菌落形成單位(CFU)方法檢測Bi-Se NPs的抗菌活性(圖2C和D)的結(jié)果顯示Bi-Se NPs組在各實(shí)驗(yàn)組中具有最顯著的抑制活性。在活/死細(xì)胞實(shí)驗(yàn)檢測中,Bi-Se NPs和20%噻菌銅處理組都顯示出了對細(xì)菌的根除效果(圖2E)。圖2F顯示Bi-Se NPs對R. solanacearum的細(xì)胞膜具有很強(qiáng)的破壞能力,導(dǎo)致細(xì)菌出現(xiàn)更多褶皺,達(dá)到抗菌效果。

圖2. Bi-Se NPs的體外抗菌特性
3.Bi-Se NPs抗菌機(jī)制探究
與對照組相比,單獨(dú)使用Bi NPs或Se NPs不足以引起細(xì)菌內(nèi)活性氧(ROS)爆發(fā)造成細(xì)菌損傷(圖3A-E)。經(jīng)過Bi-Se NPs處理后,細(xì)菌內(nèi)源性ROS水平顯著增加(圖3F)。
細(xì)菌能夠在蓋玻片表面形成致密的生物膜以避免抗菌藥物的滲透和清除。掃描電子顯微鏡(SEM)圖像證實(shí)經(jīng)過Bi-Se NPs處理后,R. solanacearum的生物膜密度顯著降低,面積較小,且抗菌生物膜活性似乎與ROS濃度呈正相關(guān),受到更強(qiáng)的破壞(圖3G和H)。與空白組相比,Bi-Se NPs處理組在590 nm處的胞外多糖(EPS)含量在各處理組中降低幅度最大(圖3I)。
Bi-Se NPs顯著增加了R. solanacearum中的ROS含量,可能影響酶、轉(zhuǎn)運(yùn)蛋白等物質(zhì)的功能,導(dǎo)致細(xì)菌細(xì)胞代謝紊亂,減少EPS產(chǎn)生,從而抑制細(xì)菌生物膜的形成,達(dá)到抑制細(xì)菌的效果。
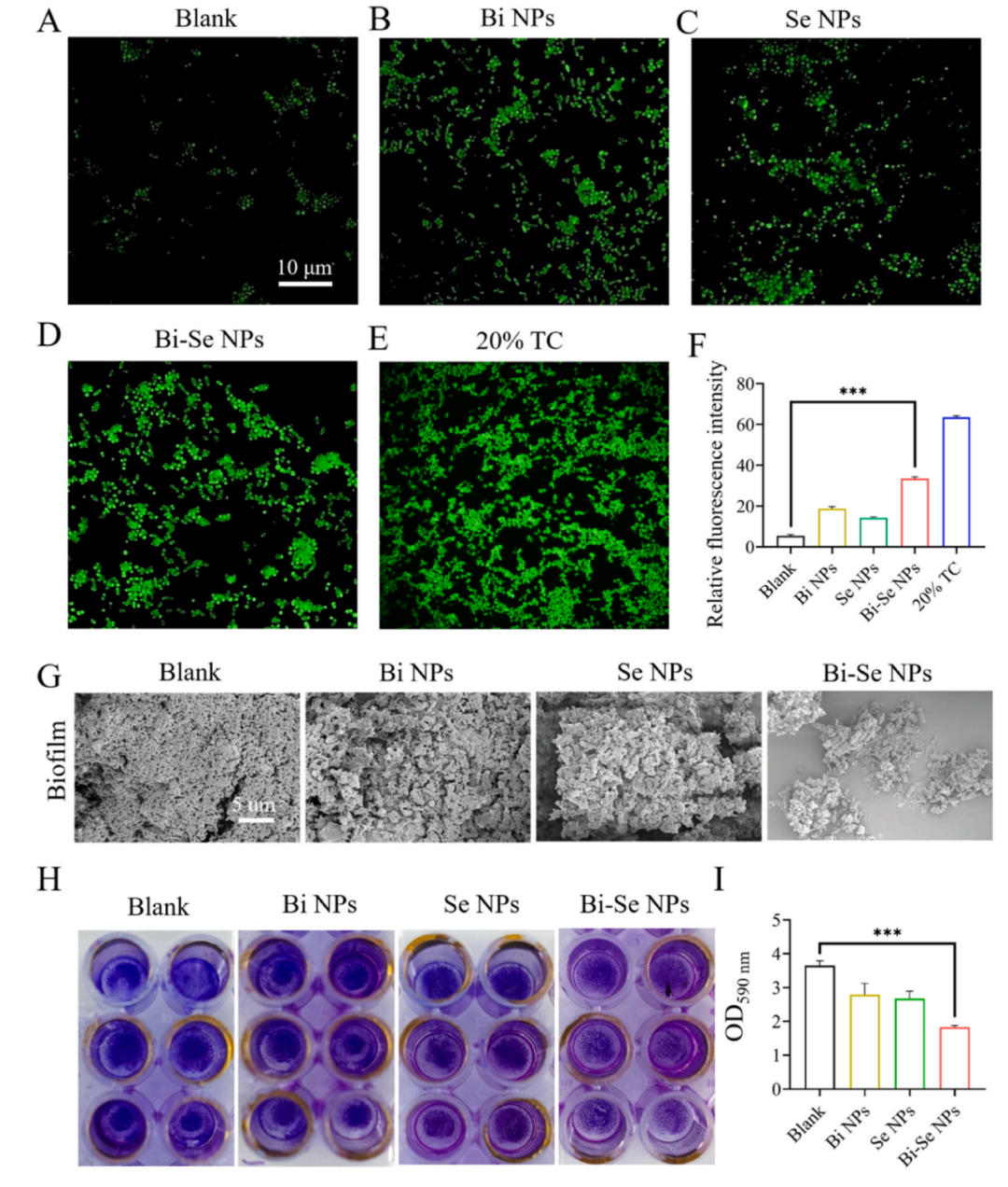
圖3. Bi-Se NPs誘導(dǎo)的活性氧(ROS)爆發(fā)和抗生物膜特性
4.不同處理下煙草的生長和理化分析
根部灌溉施用128 ± 5 μg/mL的Bi-Se NPs對添加病原體的煙草生長影響顯著(圖4A)。煙草植物吸收了Bi-Se NPs,植物高度和葉片數(shù)量增加,平均葉寬、莖圍和地上部鮮重也相應(yīng)增加,發(fā)病率指數(shù)顯著降低(圖4B-G)。煙草葉片圖像顯示(圖4H和I),經(jīng)Bi-Se NPs處理組煙草的病變較小,腐爛損傷較輕,有助于煙草的更好生長。
Bi-Se NPs通過降低煙草植物中H2O2的含量和超氧化物歧化酶(SOD)的活性,減少其對植物造成的損害(圖5A和B)。此外,原糖和總糖的含量增加的同時(shí)降低總堿含量,有助于積累營養(yǎng)物質(zhì)并支持煙草植物的生長和發(fā)育(圖5C-E)。Bi-Se NPs的添加還顯著增加了煙草葉片中的硒含量(圖5F)。經(jīng)處理組的葉片葉表面褶皺較少,葉脈更粗,葉片展開度更大,生長更為有利(圖5G)。
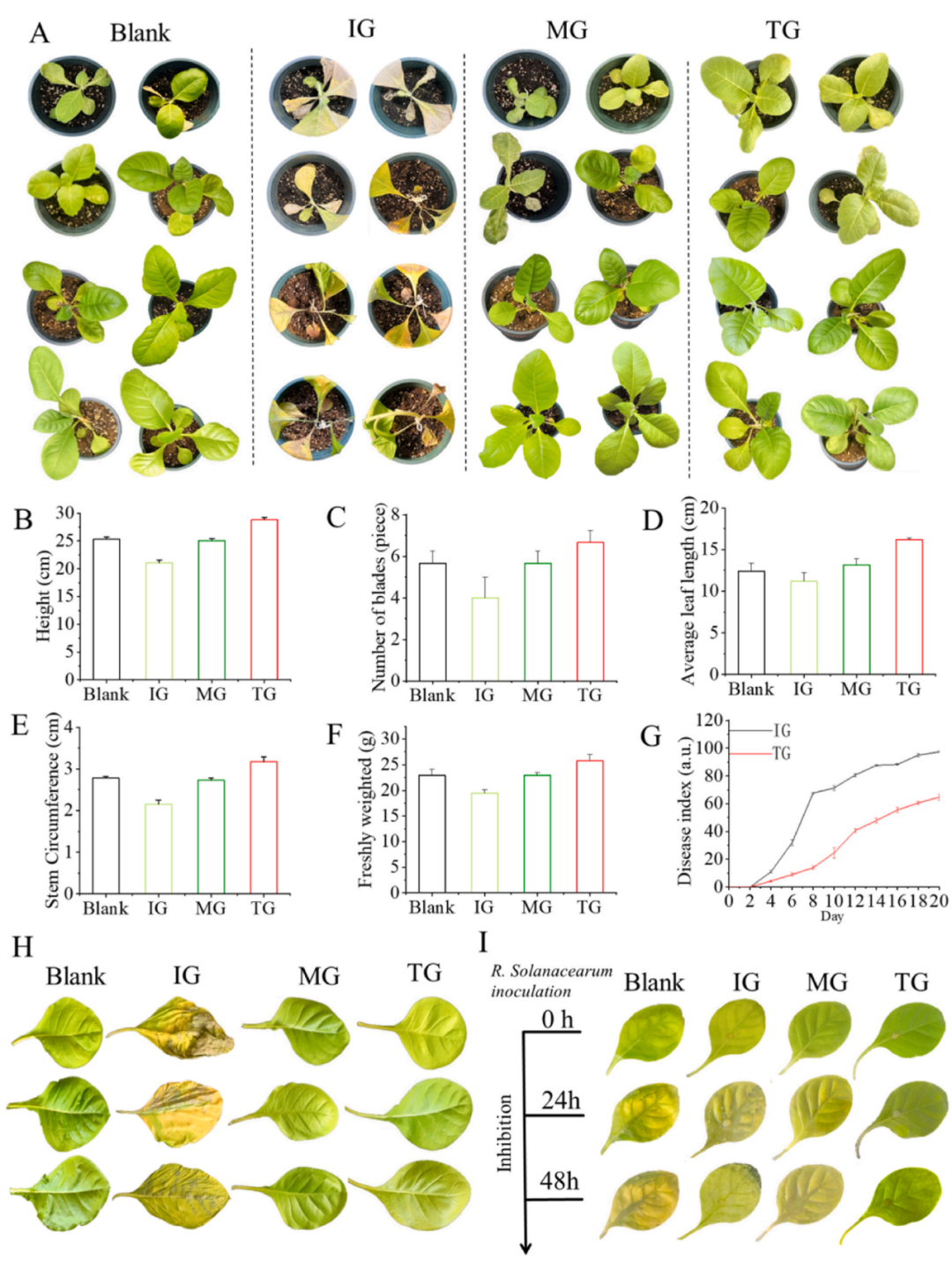
圖4. 施用128 ± 5 μg/mL Bi-Se NPs后煙草幼苗的生長
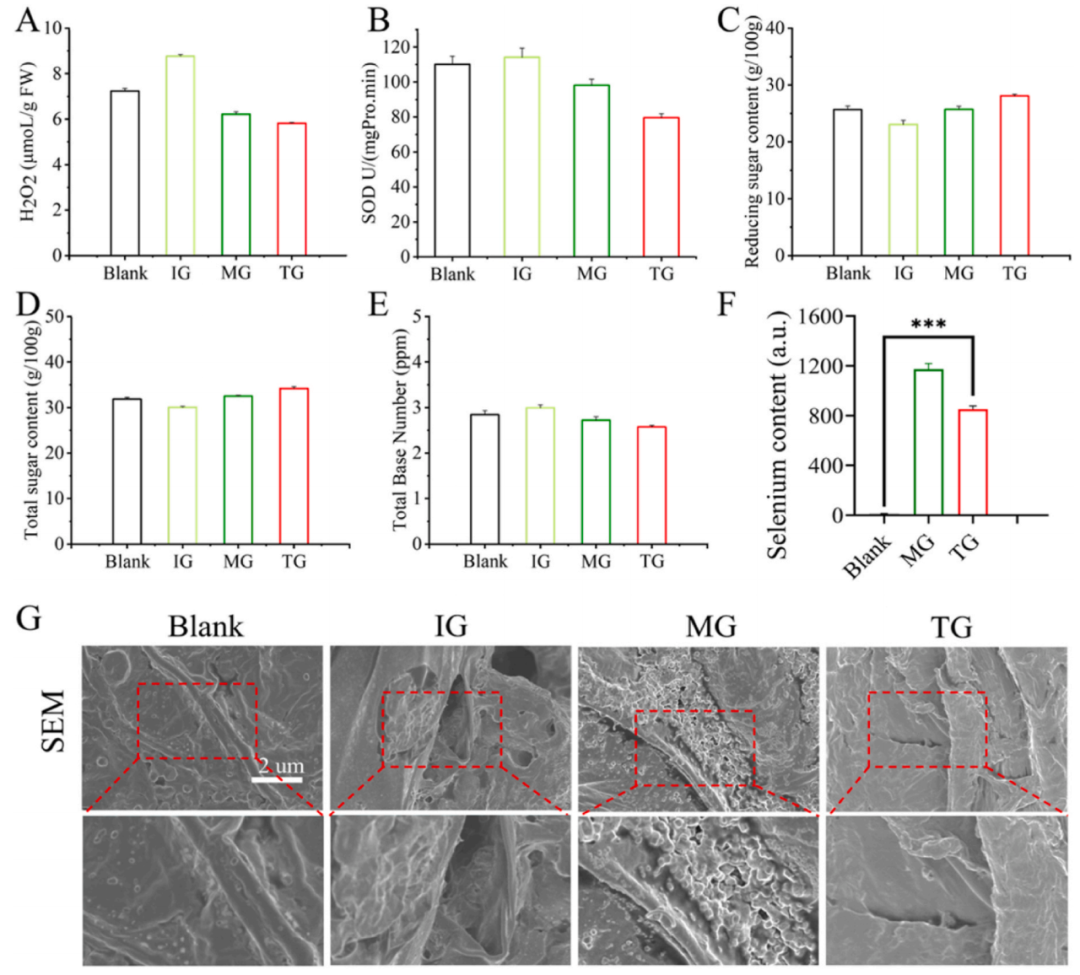
圖5. 煙草葉片的生理生化檢測和形態(tài)學(xué)分析
5.不同處理下煙草非靶向代謝組的差異表達(dá)分析
正負(fù)離子結(jié)合代謝物鑒定發(fā)現(xiàn),羧酸及其衍生物的化學(xué)分類和歸屬信息約占所有鑒定差異代謝物的13.4%(圖6A)。熱圖分析結(jié)果顯示不同處理的樣本之間代謝物存在較大差異(圖6B)。火山圖分析樣本之間的差異代謝物(圖6C)發(fā)現(xiàn)約有240種差異代謝物上調(diào),約115種差異代謝物下調(diào)。顯著差異代謝物之間的相關(guān)性緊密程度分析結(jié)果顯示,羧酸及其衍生代謝物琥珀酸變異系數(shù)約為3倍(圖6D和E)。利用KEGG數(shù)據(jù)庫對差異代謝物的途徑進(jìn)行注釋,并獲得差異代謝物的代謝途徑圖,其中氧化磷酸化途徑富集程度最高(圖6F)。進(jìn)一步分析氧化磷酸化途徑(圖6G),發(fā)現(xiàn)琥珀酸參與了氧化磷酸化途徑的運(yùn)行,加速煙草中氧氣和ATP的產(chǎn)生,從而促進(jìn)機(jī)體代謝。
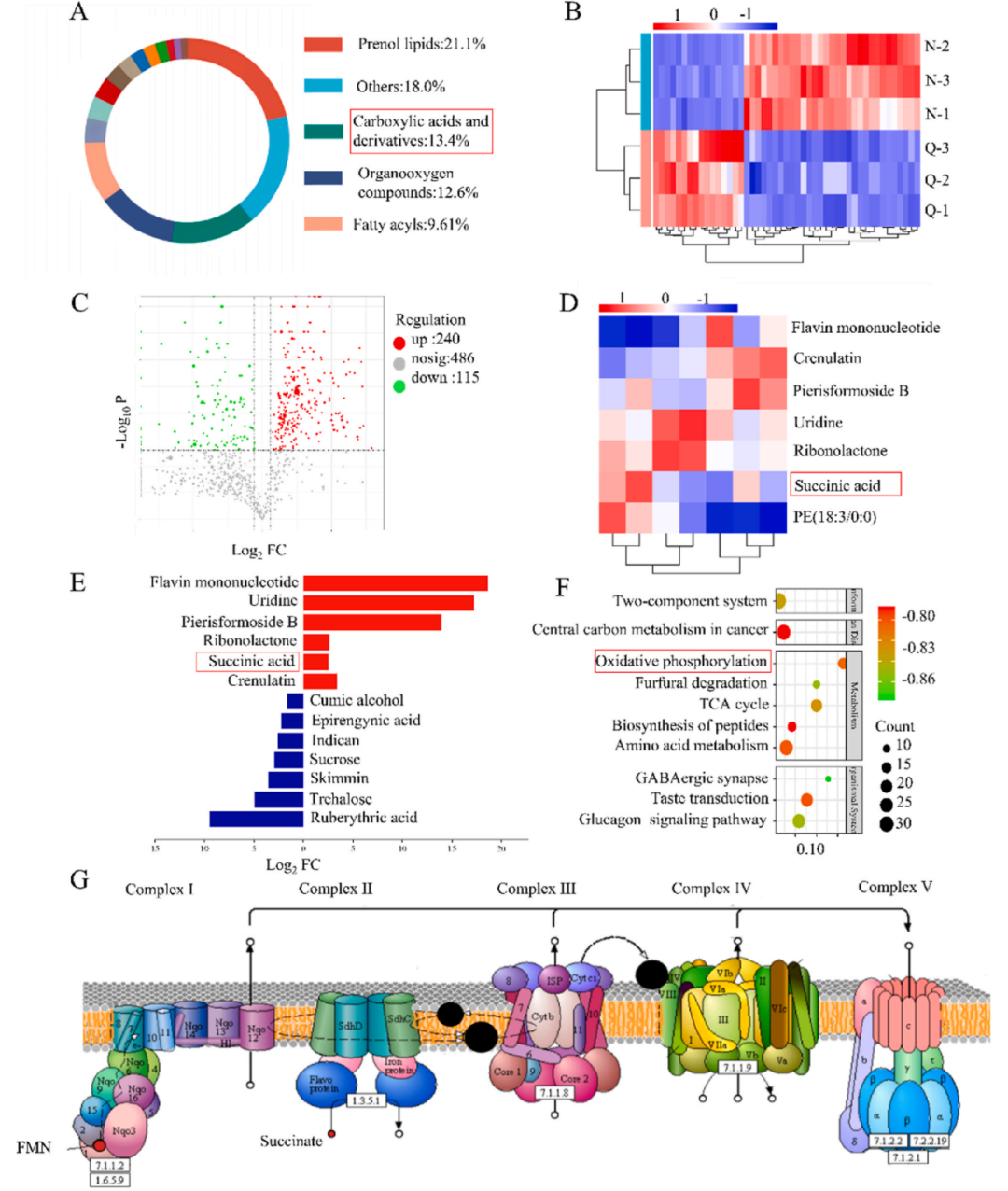
圖6. 不同處理組煙草葉片的非靶向代謝組檢測
| 三黍服務(wù) | 技術(shù)儀器 |
非靶代謝組學(xué)(LC-MS) | LC-MS(Q Exactive HF-X) |
非靶代謝組學(xué)(GC-MS) | GC-MS(Agilent 7820,5977) |
植物廣泛靶向代謝組學(xué) | |
風(fēng)味組學(xué) | |
中藥代謝組學(xué) | |
更多服務(wù),歡迎私信詳詢! | |
6.不同處理下煙草真核轉(zhuǎn)錄組的差異表達(dá)分析
轉(zhuǎn)錄組學(xué)分析顯示Bi-Se NPs的存在可以提高煙草中氧化磷酸化途徑的運(yùn)行效率,從而促進(jìn)煙草的生長和發(fā)育。
火山圖分析直觀地顯示了樣本之間差異基因的變化,其中約有4239個(gè)差異調(diào)控基因上調(diào),約5308個(gè)差異調(diào)控基因下調(diào),并對檢測到的差異基因進(jìn)行熱圖分析,RT-qPCR驗(yàn)證三個(gè)上調(diào)基因和一個(gè)下調(diào)基因結(jié)果與轉(zhuǎn)錄結(jié)果一致(圖7A到圖7C)。通過GO注釋和KEGG數(shù)據(jù)庫對差異基因進(jìn)行分析,發(fā)現(xiàn)氧化磷酸化途徑的富集程度最高(圖7D和E)。
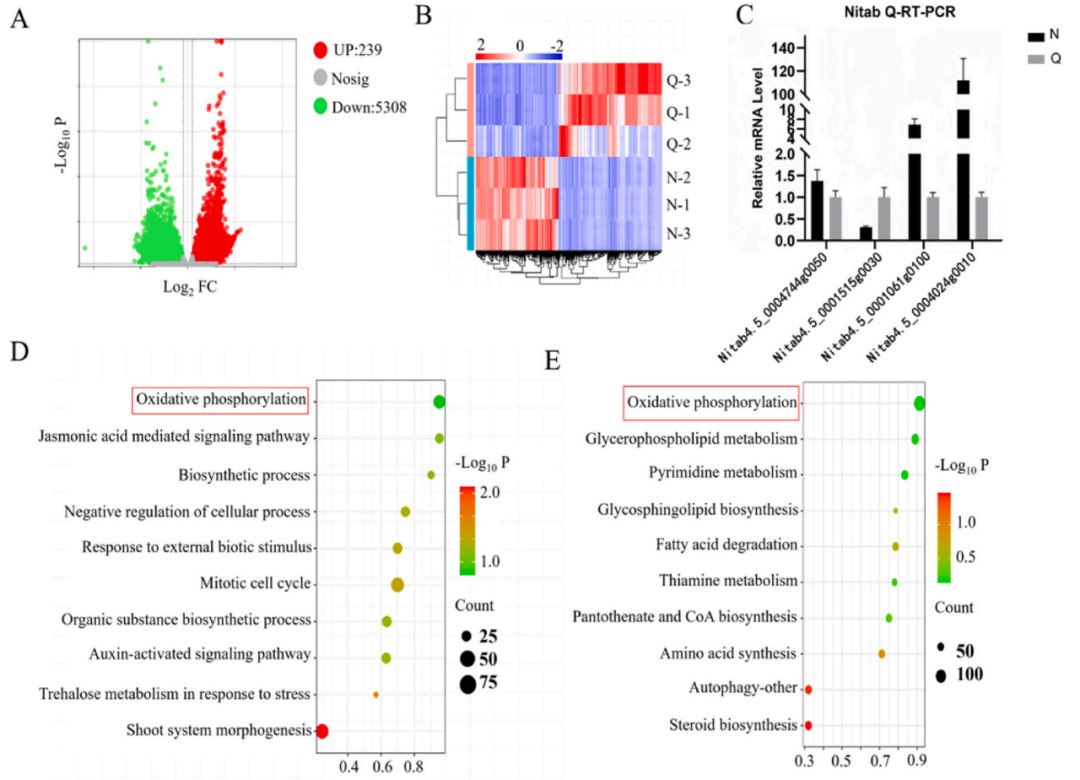
圖7. 不同處理組煙草葉片的真核轉(zhuǎn)錄組檢測
| 三黍服務(wù) | 技術(shù)儀器 |
真核轉(zhuǎn)錄組測序(有參/無參) | Illumina/ BGI T7 PE150(6G) |
原核轉(zhuǎn)錄組測序 | Illumina PE150(2G) |
全長擴(kuò)增子測序 | |
DAP-seq測序 | |
10×單細(xì)胞轉(zhuǎn)錄組測序(3') | |
更多服務(wù),歡迎私信詳詢! | |
7.不同處理下煙草磷酸化蛋白質(zhì)組差異表達(dá)分析
不同處理下的煙草生長差異導(dǎo)致了差異蛋白的出現(xiàn)。樣本之間差異蛋白的火山圖分析(圖8A)直觀地顯示了差異蛋白的變化,其中約有656個(gè)差異調(diào)控蛋白上調(diào),約320個(gè)差異蛋白下調(diào)。對檢測到的差異蛋白進(jìn)行熱圖分析結(jié)果如圖8B所示。分析差異蛋白在GO注釋中的分布,與背景蛋白相比,代謝過程的調(diào)控高度富集,約為51(圖8C)。對差異蛋白進(jìn)行KEGG富集分析,其中包含的代謝途徑和氧化磷酸化途徑高度富集,表明Bi-Se NPs的存在提高了這些途徑的運(yùn)行效率(圖8D)。最后,分析了差異蛋白之間的蛋白質(zhì)-蛋白質(zhì)相互作用網(wǎng)絡(luò)(圖8E),其中與LOC107795313節(jié)點(diǎn)對應(yīng)的核孔蛋白與其他蛋白的相互作用最為密切,表明該蛋白與煙草的生長發(fā)育過程密切相關(guān),進(jìn)一步驗(yàn)證了Bi-Se NPs對煙草代謝的影響。
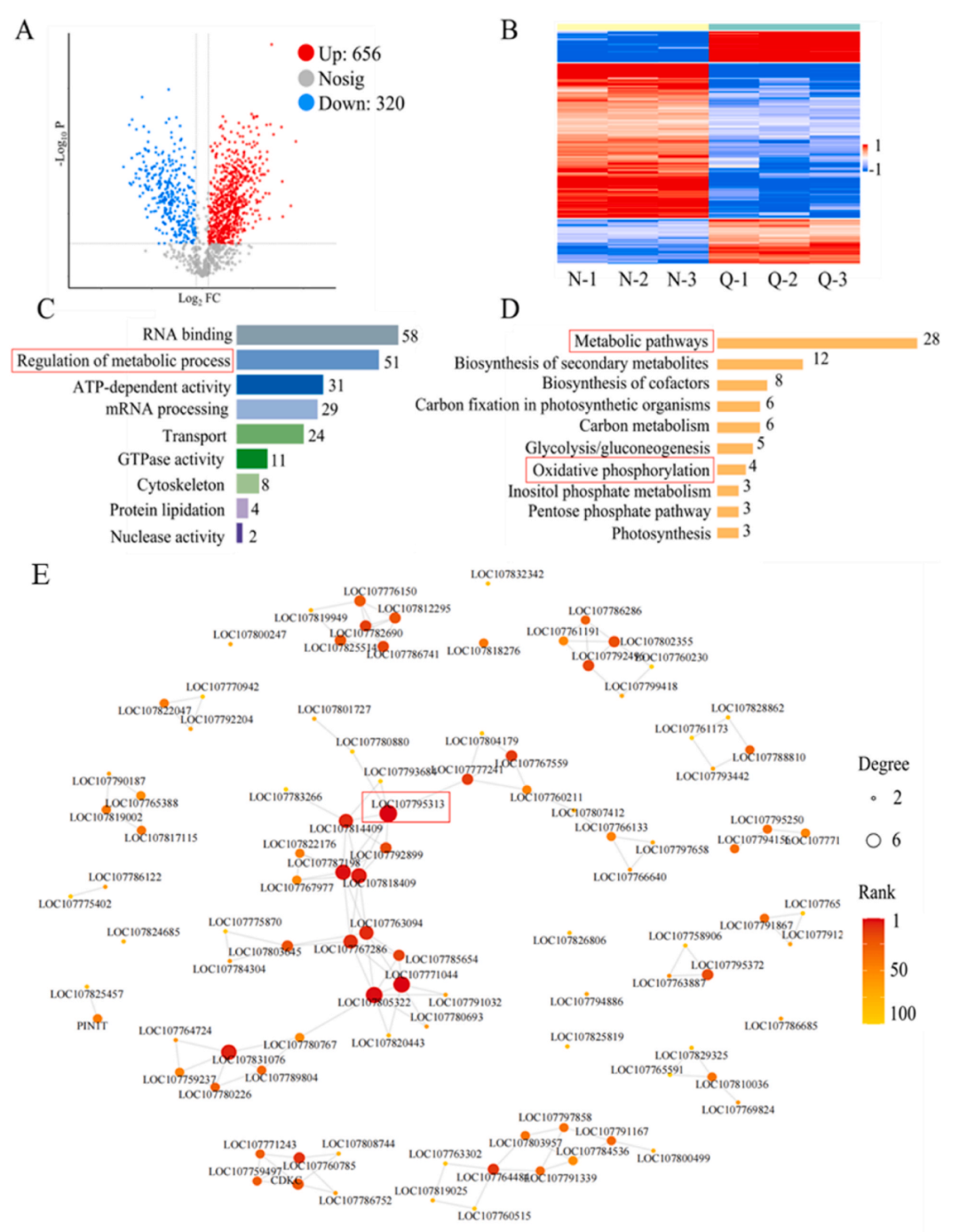
圖8. 不同處理組煙草葉片的磷酸化蛋白質(zhì)組檢測
| 三黍服務(wù) | 技術(shù)儀器 |
Astral DIA 磷酸化修飾蛋白組分析 | |
4D-DIA 泛素化修飾蛋白組分析 | |
4D-DIA 乙酰化修飾蛋白組分析 | |
N-糖基化蛋白質(zhì)組學(xué)完整糖肽分析 | |
N-糖基化蛋白質(zhì)組學(xué)位點(diǎn)鑒定分析 | |
O-糖基化蛋白質(zhì)組學(xué)完整糖肽分析 | |
更多服務(wù),歡迎私信詳詢! | |
8.不同處理下煙草多組學(xué)分析
綜合代謝組學(xué)、轉(zhuǎn)錄組學(xué)和蛋白質(zhì)組學(xué)數(shù)據(jù),研究團(tuán)隊(duì)發(fā)現(xiàn)Bi-Se NPs通過多種途徑抗菌并促進(jìn)煙草生長發(fā)育。
轉(zhuǎn)錄組與蛋白質(zhì)組聯(lián)合分析韋恩圖顯示了基因和蛋白質(zhì)之間定量和差異表達(dá)的信息,從而得出差異基因和差異蛋白之間的對應(yīng)關(guān)系(圖9A)。此外,研究者還根據(jù)每個(gè)對齊組中蛋白質(zhì)表達(dá)的倍數(shù)差異和基因表達(dá)的倍數(shù)差異計(jì)算相關(guān)系數(shù),并通過聚類分析可視化了所有可定量的蛋白質(zhì)和基因之間的關(guān)聯(lián)(圖9B和C)。不同樣本之間差異基因和差異蛋白存在一定程度的差異。
根據(jù)上述數(shù)據(jù),GO途徑在兩種組學(xué)中均顯著富集(圖9D),其中差異蛋白在代謝過程調(diào)控中的富集更為顯著,差異基因在色素代謝中的富集最為顯著。KEGG途徑在兩種組學(xué)數(shù)據(jù)中均顯著富集(圖9E),表明植物激素信號轉(zhuǎn)導(dǎo)、泛酸和輔酶A生物合成以及光合作用生物中的碳固定在差異蛋白或差異基因中發(fā)生了更顯著的變化。
從差異基因和差異蛋白的角度來看,施用128 ± 5 μg/mL的Bi-Se NPs后,植物激素信號轉(zhuǎn)導(dǎo)、泛酸和輔酶A生物合成以及光合作用生物中的碳固定含量顯著增加,促進(jìn)了煙草體內(nèi)代謝調(diào)節(jié),提高了氧化磷酸化途徑和光合作用的運(yùn)行效率,增加了激素信號轉(zhuǎn)導(dǎo)的含量,并提高了琥珀酸和FMN的水平。在生物合成方面,Bi-Se NPs施用促進(jìn)了煙草的代謝和生長發(fā)育能力,抑制了R. solanacearum的活性,降低了煙草青枯病的發(fā)病率,表明Bi-Se NPs可以保護(hù)煙草免受病原細(xì)菌感染,并促進(jìn)植物的生長發(fā)育。
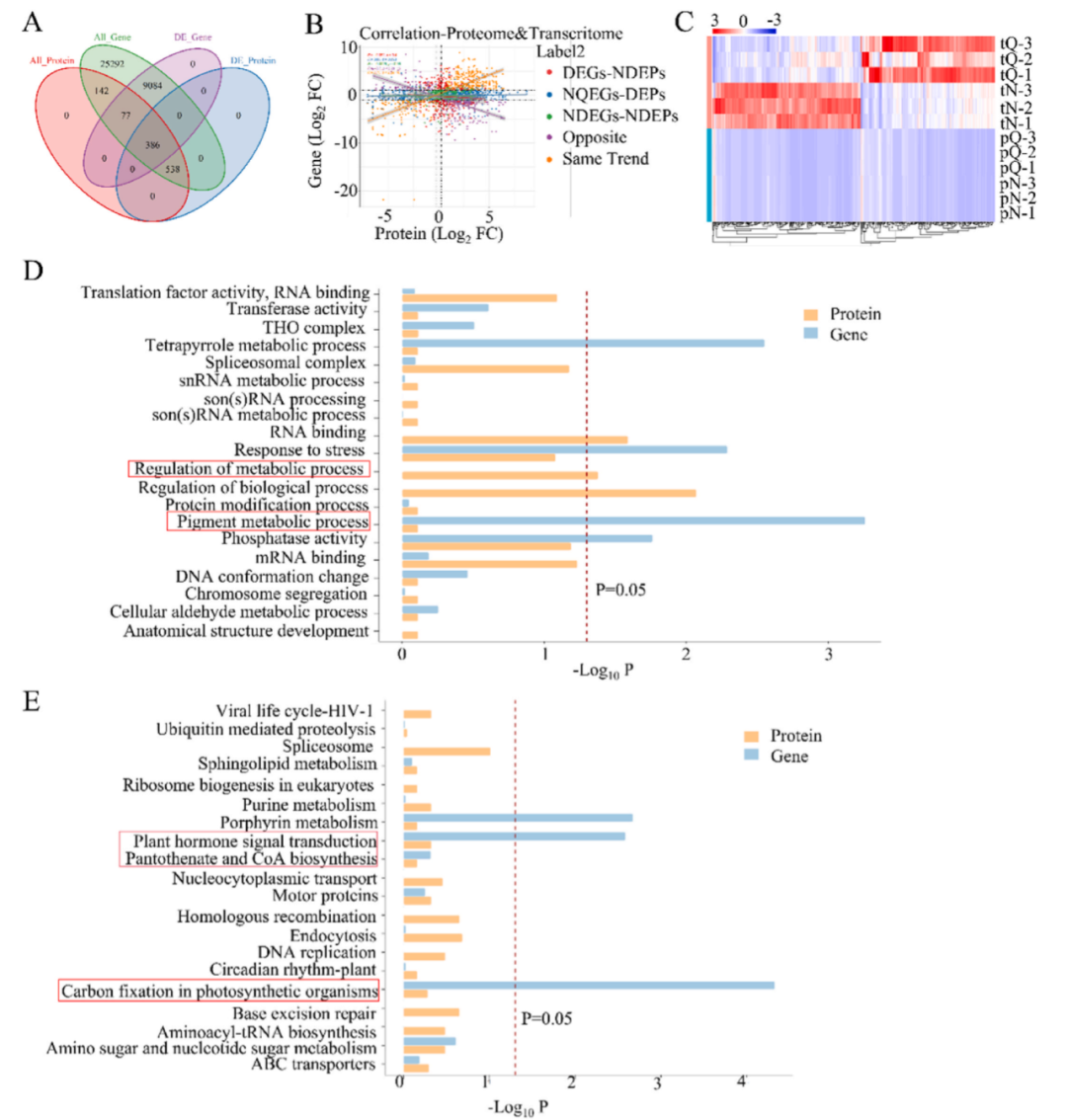
圖9. 不同處理組煙草葉片的多組學(xué)分析
9.安全性評估
此外,作者還針對Bi-Se NPs的安全性進(jìn)行了評估。與對照組相比,使用Bi-Se NPs對心肌細(xì)胞的正常生長沒有顯著影響。對小鼠進(jìn)行的Bi-Se NPs處理也表現(xiàn)出Bi-Se NPs具備低毒性,具有良好的安全性和可靠性。
除了對小鼠的安全性測試外,還對土壤環(huán)境中微生物的Bi-Se NPs安全性進(jìn)行了檢查。抑菌圈實(shí)驗(yàn)表明合理使用適當(dāng)濃度的Bi-Se NPs具有良好的生物和環(huán)境安全性。
本研究通過合成新型的鉍-硒納米復(fù)合簇(Bi-Se NPs),成功探索了其對煙草青枯病的防治效果及其作用機(jī)制。Bi-Se NPs不僅具有良好的抗菌性能,還能顯著促進(jìn)煙草的生長和發(fā)育,提高植物的抗逆性。通過多組學(xué)分析,研究團(tuán)隊(duì)揭示了Bi-Se NPs在植物代謝調(diào)節(jié)中的多重作用機(jī)制,為納米材料在農(nóng)業(yè)領(lǐng)域的應(yīng)用提供了新的視角。此外,Bi-Se NPs在生物安全性和環(huán)境安全性方面的良好表現(xiàn),為其在實(shí)際農(nóng)業(yè)生產(chǎn)和植物病害防治中的應(yīng)用奠定了堅(jiān)實(shí)的基礎(chǔ)。
滑動(dòng)查看更多:


排版:野凌
審核:三黍生物企宣部

