










【背景】
油莎草(學名:Cyperus esculentus L.)是莎[suō]草科莎草屬植物。它含有許多成分,包括淀粉、蛋白質、脂肪、生物堿、酚類和類黃酮等,已被廣泛應用于食品加工、生物醫藥和生物燃料開發等領域。文獻也證實了其保護胃腸道、降血糖等特性。
乙酰化、磺化、羧甲基化和磷酸化常被用于修飾多糖結構,某些官能團的引入改變了分子量和其他結構特征,從而影響了生物功能特性。其中乙酰化可拉伸多糖分支,改變多糖鏈的空間排列,使羥基暴露,從而改變水溶性和疏水性。然而,改性對油莎草結構和性質的影響尚不清楚。
本文研究了乙酰化對油莎草的結構特征、理化性質、熱穩定性、抗氧化和免疫調節活性的影響。

【樣本類型】
油莎草多糖
【技術方法】
可溶性糖檢測,總蛋白含量測定,還原糖檢測,乙醛酸檢測,紫外光譜掃描、紅外光譜掃描、熱重、掃描電鏡、甲基化、核磁掃描、抗氧化活性檢測(DPPH自由基清除活性測定、羥自由基清除活性測定、ABTS自由基清除活性測定、鐵還原抗氧化能力測定、超氧陰離子清除活性測定),細胞實驗:抗炎能力測定、RAW264.7細胞損傷修復能力、ROS分泌促進能力等。
【研究結果】
1、 乙酰化對油莎草多糖(CEP)的化學成分影響
乙酰化油莎草多糖(ACEP)的總糖、蛋白質、還原糖和乙醛酸水平顯著低于CEP (P< 0.05)。CEP可能在改性過程中被降解,導致改性后各種化合物的量大幅減少。這些結果與之前的報告相似。
乙酰化前CEP表面粗糙,有凹陷、不均勻和粘附。乙酰化后,多糖顆粒破碎明顯,呈不規則片狀和短棒狀,結構松散。這可能是由于CEP大多以聚集狀態發生,具有較強的分子間交聯,而乙酰化后分子間力降低,相關基團被釋放。掃描電鏡觀察的多糖顆粒的表面形態是由分子間交聯引起的各種分子聚集引起的,這種聚集引起構象變化,隨后產生獨特的形態特征。

圖1 CEP(A1,500 ×;A2, 1000x;A3, 2000 x;A4,5000 × )和ACEP(A1,500 ×;A2, 1000x;A3, 2000 x;A4,5000 × )的掃描電鏡圖片
2、乙酰化對CEP性質的影響
CEP和ACEP的分子量分別為148和138 kDa,ACEP分子量的降低可能與多糖的降解有關(如圖2A和2B)。CEP和ACEP是由葡萄糖、甘露糖、阿拉伯糖和木糖組成的雜多糖,而乙酰化并沒有顯著改變單糖的組成,而只是影響了比例(如圖2C)。乙酰化可能導致乙酰化衍生物的分子量降低,但不會破壞多糖的主鏈。
CEP和ACEP在260 ~ 280 nm處有較弱的吸收峰,說明存在微量蛋白(如圖2D)。與CEP的FTIR結果相比(圖2E), ACEP在1720 cm-1和1247 cm-1處有很強的吸收峰。這些分別代表乙酰酯基團C-O和C-O拉伸振動,表明乙酰基修飾成功。
乙酰化對CEP熱穩定性的影響該實驗證明了糖的穩定性(如圖2F)。CEP和ACEP有三個減重階段:30°C - 200°C、200°C -375°C和375°C-800°C。在第二階段,ACEP降解速度更快,表明其具有更好的熱降解性能。但隨著溫度的升高,ACEP的重量變化相對緩慢,800℃時的剩余重量(12.46%)大于CEP(10.28%)。結果表明,乙酰化處理顯著提高了CEP的熱穩定性。
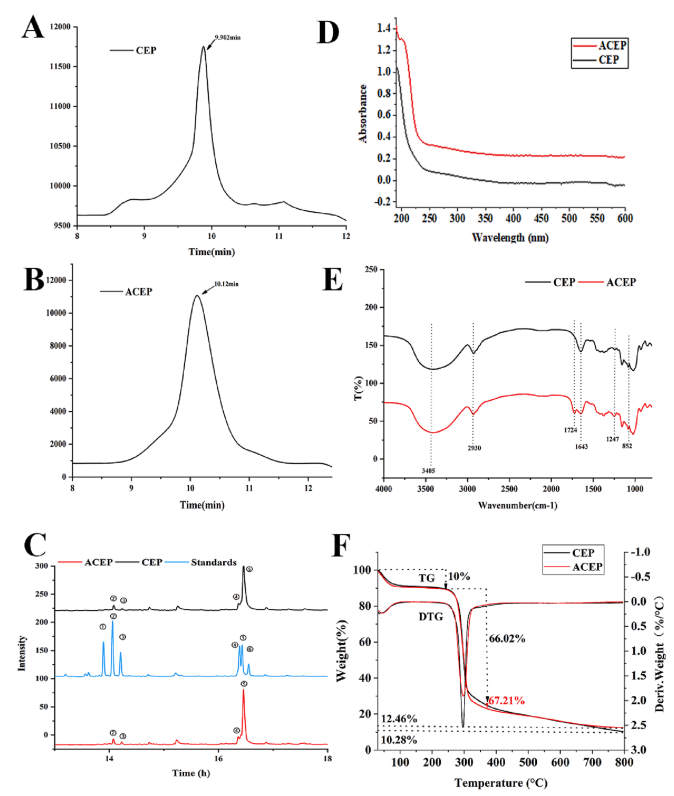
圖2 (A、B) CEP和ACEP的HPLC洗脫圖譜。(C)標準單糖、CEP和ACEP(-鼠李糖、-阿拉伯糖、3-木糖、④-甘露糖,-葡萄糖,-半乳糖)(D) CEP和ACEP的紫外光譜。(E) CEP和ACEP的FTIR光譜。(F) CEP和ACEP的TGA。
3、 乙酰化對CEP抗氧化能力的影響
CEP和ACEP均能有效清除DPPH自由基(如圖A),且乙酰化能增強CEP的清除能力。與DPPH結果相似,乙酰化提高了CEP對ABTS自由基(如圖B)、對OH自由基(如圖C)、FRAP自由基(如圖D) 、超氧陰離子自由基(如圖E)的清除能力。綜上所述,乙酰化多糖的抗氧化活性優于天然乙酰化多糖。
植物多糖清除自由基的能力受其分子結構和理化性質的影響。乙酰化修飾可以改變多糖的理化性質,從而改善其水溶性和生物活性。先前的研究表明,乙酰基可能取代多糖的C-2和/或C-3,而雜資本碳(heterocapital carbon)的氫原子活化增加了所得衍生物的供氫能力。在本研究中,ACEP的抗氧化能力超過CEP,這可能是由于引入了乙酰基以提高供氫能力。在CEP的C-2和C-6位置上取代的乙酰基可以激活端粒碳的氫原子。除了乙酰基本身外,多糖的組成和相對分子質量也與多糖的生物活性有關。低分子量多糖可以取代更多的羥基末端來消除自由基。改性后的CEP分子量降低,可能是其具有較強的自由基清除能力的原因。多項研究表明,化學修飾可以通過改變多糖的組成來提高多糖的抗氧化活性, Ara和Gal含量較高的多糖抗氧化活性更高。SEM結果也證實了ACEP的表面更加展開,這增加了其在溶液中的接觸面積,有助于提高ACEP的自由基清除率,從而賦予其優越的抗氧化能力。
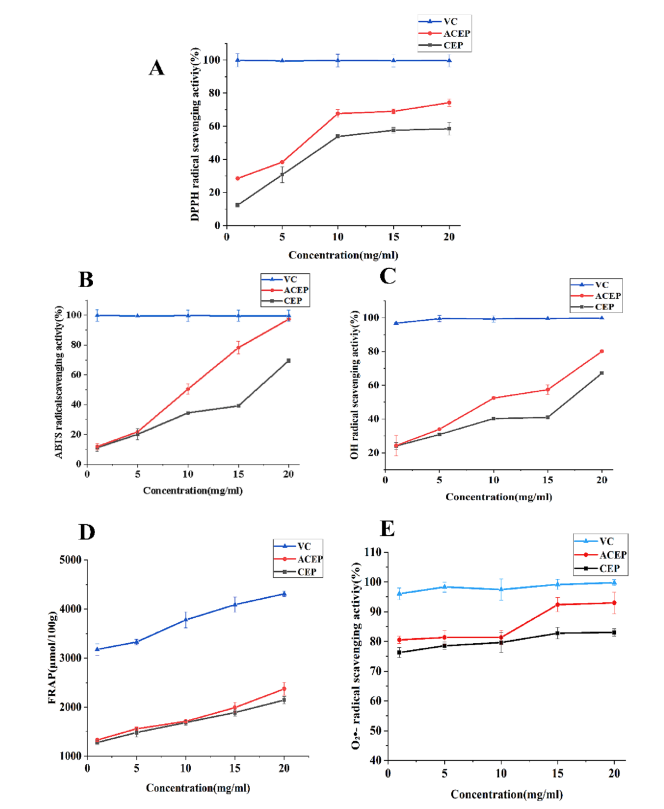
圖3 (A) CEP和ACEP對DPPH自由基的清除活性。(B) CEP和ACEP對ABTS自由基的清除活性。(C) CEP和ACEP對eOH自由基的清除活性。(D) CEP和ACEP對FRAP自由基的清除活性。(E) CEP和ACEP對眼自由基的清除活性。數值表示為平均值±SD (n = 3)。
4、 乙酰化對lps誘導RAW264.7細胞CEP抗炎活性的影響
如圖4A所示,CEP和ACEP對RAW 264.7細胞存活率無顯著影響(P > 0.05)。
如圖4B所示,與對照組相比,LPS顯著降低了RAW 264.7細胞的活力(P < 0.05)。與CEP和ACEP孵育后,RAW 264.7細胞存活率從CEP的76%提高到86%,ACEP的93%,顯著提高(P< 0.05)。說明CEP和ACEP均能有效減輕LPS對RAW264.7細胞的免疫損傷,且ACEP的免疫保護作用強于CEP。
細胞形態倒置顯微鏡下觀察不同濃度CEP和ACEP(125、500和1000 μg/mL)對lps誘導的RAW 264.7細胞形態的影響。如圖4C所示,對照組細胞呈圓形,生長正常。LPS處理后,細胞形態由橢圓變為伸長,大部分細胞開始漏漿。CEP或ACEP預孵育后,細胞形態有恢復趨勢,隨著濃度的增加,拉伸細胞數量減少,細胞形態與對照組更接近(圖片空白越多表明細胞受破壞程度越嚴重)。結果表明,CEP和ACEP都能夠修復lps誘導的RAW264.7細胞損傷,其中ACEP的修復能力更強。
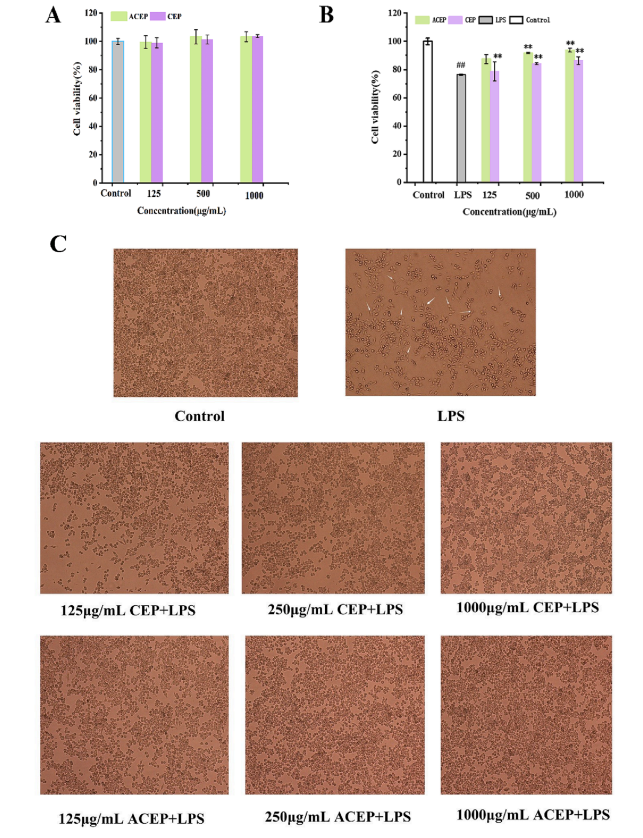
圖4 (A) CEP和ACEP對RAW264.7細胞活力的影響。(B) CEP和ACEP對lps誘導的RAW264.7細胞活力的影響。(C) CEP和ACEP對lps誘導的RAW 264.7細胞形態的影響。Scar-100微米。用不同濃度的CEP或ACEP預處理細胞12 h,然后暴露于LPS (1 ug/ml)中12 h。倒置光學顯微鏡下觀察細胞形態。數值表示為平均值±SD (n=6)。“P< 0.05”,“P< 0.001”vs對照組。與LPS組比較,P< 0.05, **P< 0.001。
ROS分泌ROS是參與正常生理代謝活動不可缺少的信號分子。巨噬細胞產生過多的ROS,刺激免疫反應(Chu et al., 2013)。如圖5所示,LPS (1 μg/mL)刺激12 h后,DCFH-DA熒光強度顯著增強,大部分細胞由圓形變為拉伸形態,表明ROS濃度顯著升高。但不同濃度CEP和ACEP(125、500、1000 μg/mL)孵育細胞時,熒光強度呈劑量依賴性逐漸降低,且ACEP組熒光強度低于CEP組。上述結果表明,CEP和ACEP均能顯著抑制lps誘導的RAW264.7細胞中ROS的產生,其中乙酰化多糖的作用更大。
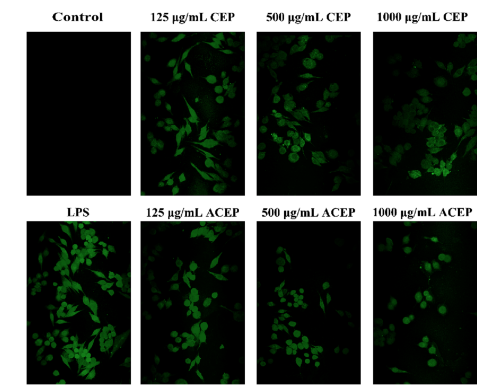
圖5 CEP或ACEP抑制LPS中ROS的生成。將細胞分別用不同濃度的CED或ACEP處理12 h,然后用LPS (1 ug/mL)處理12 h,共聚焦激光掃描顯微鏡觀察細胞的變化。細胞的熒光顯微鏡圖像表示ROS。
【小結】
在這項研究中,作者對油莎草多糖及其乙酰化多糖的化學性質、抗氧化特性和免疫調節活性進行了研究。結果表明,CEP和ACEP是由葡萄糖、甘露糖、阿拉伯糖和木糖組成的雜多糖。CEP的主鏈包括a-1,4- glp殘基,分支點位于a-1,6- manp殘基的O-6位置。在一些葡萄糖殘基的O-2和O-6位置上取代了乙酰基。乙酰化顯著改善了ACEP的熱穩定性,并表現出較好的抗氧化活性。CEP和ACEP均能有效緩解和修復RAW264.7細胞的免疫損傷,抑制ROS的產生。ACEP的抗炎作用也比CEP更強。綜上所述,CEP和ACEP均可作為潛在的天然抗氧化劑或免疫調節劑應用于食品科學和生物學。
【往期推薦】
文獻解讀 | 大規模基因組和轉錄組學分析闡明雞高產的遺傳基礎
文獻解讀 | 如何進行中藥復方的體內機制研究,學會這個思路就夠了!


