【背景】
雞是生產(chǎn)蛋白質(zhì)效率最高的動(dòng)物之一,為人類提供了超過30%的肉制品,在全球糧食安全中發(fā)揮著重要作用。雖然世界上有數(shù)百個(gè)家養(yǎng)雞品種,但其肉類產(chǎn)量低,無法滿足人類的需求。快速生長的白羽肉雞被培育出來,通過對生產(chǎn)性狀的集約化選擇,雞的生產(chǎn)力不斷提高。此外,白羽肉雞的飼料轉(zhuǎn)化率是牛的10倍,但其碳排放量僅為牛的1/10。這與肉雞的高產(chǎn)肉量密切相關(guān)。然而,通過集約化選擇培育出的純種肉雞肉制品的遺傳基礎(chǔ)尚未得到充分研究。
2023年3月發(fā)表在Journal of Advanced Research上的題為“Large-scale genomic and transcriptomic analyses elucidate the genetic basis of high meat yield in chickens”的論文,對3個(gè)純種肉雞(n=748)和6個(gè)地方品種/系(n=114)進(jìn)行了全基因組重測序,并從NCBI數(shù)據(jù)庫獲得了12個(gè)雞品種(n=199)的測序數(shù)據(jù)。此外,還對兩個(gè)雞品種(n=129)兩個(gè)發(fā)育階段的六種組織進(jìn)行了轉(zhuǎn)錄組測序。隨后采用基因組關(guān)聯(lián)分析結(jié)合cis-eQTL定位和Mendel隨機(jī)化方法進(jìn)行了分析。
【樣本類型】
3個(gè)純種肉雞(n=748),6個(gè)地方品種/系(n=114)
【技術(shù)方法】
全基因組測序,轉(zhuǎn)錄組測序,全基因組關(guān)聯(lián)分析,cis-eQTL定位和Mendel隨機(jī)化方法。
【圖形摘要】

【研究結(jié)果】
全基因組測序和變異
本研究共獲得了862只雞的全基因組序列,共7.7 Tb原始數(shù)據(jù),平均深度為9.2倍,其余199只雞的其他序列來自SRA數(shù)據(jù)庫。采用嚴(yán)格的變異過濾后,在21個(gè)雞品種/品系中共檢測到17622141個(gè)高質(zhì)量SNP。保留的SNP分布在整個(gè)基因組中,平均密度為每60個(gè)堿基1個(gè)SNP。經(jīng)標(biāo)注后,GGA16上的新SNP與已知SNP的比例最高,其中21.74%的SNP未被包含在dbSNP中(圖1B)。B系是長勢快速的純種肉雞,羽毛呈白色,攜帶所有變異SNP 8,663,580個(gè),平均密度為每114個(gè)堿基中有1個(gè)SNP。其中只有4.7%的SNP被鑒定為新的變異。所有品種的大多數(shù)SNP都位于內(nèi)含子和基因間區(qū)域。近幾十年來,在純種肉雞身上進(jìn)行了對特定性狀的密集選擇,導(dǎo)致遺傳多樣性的減少,包括純種肉雞SNP數(shù)量和核苷酸多樣性的降低(圖1C-D)。圖中只展示了純種肉雞、野生祖先和一些典型的生長緩慢的地方品種雞。
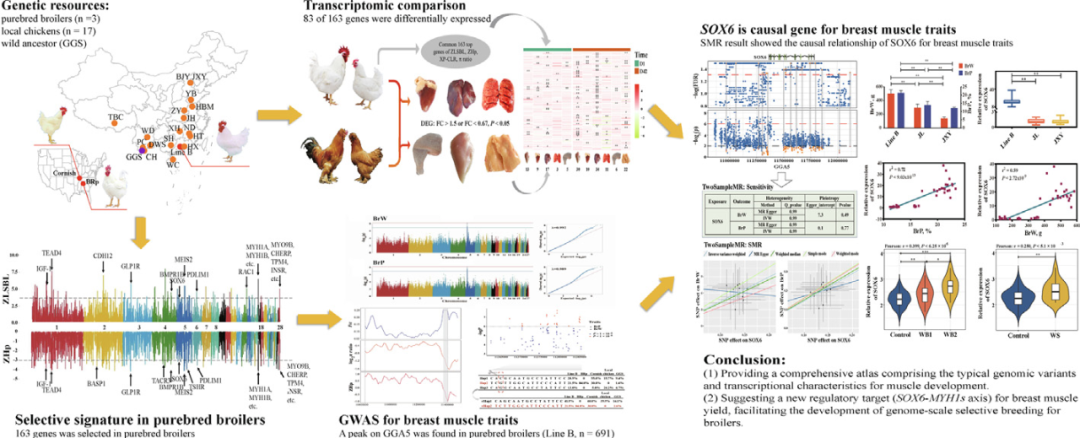
圖1 21個(gè)雞品種/系基因組信息統(tǒng)計(jì)
系統(tǒng)發(fā)育和數(shù)量統(tǒng)計(jì)學(xué)分析
本研究對這些品種間的遺傳關(guān)系進(jìn)行了綜合分析。首先,作者基于SNP對21個(gè)雞品種/品系進(jìn)行PCA(圖2A)。進(jìn)一步進(jìn)行了遺傳共起源分析,通過改變推測的祖先群體數(shù)量將所有雞劃分為不同的群體(K=2-10,圖2B)。結(jié)果發(fā)現(xiàn),品種B在基因上接近康沃爾雞品種,并且兩者都與BRp品種、本地雞品種和GGS品種分離。以GGS為外群,基于配對遺傳距離構(gòu)建鄰接樹(圖2C)。結(jié)果顯示,快速生長的純種肉雞與GGS的遺傳距離很遙遠(yuǎn),這證明了對快速生長的純種肉雞具有更強(qiáng)的選擇力,這與純種肉雞連鎖不平衡衰減的結(jié)果相一致(圖2D)。然而,純種肉雞(594-720 kb)的連續(xù)純合片段長度略低于本地肉雞(509-1012 kb)和野生祖先(1198 kb)(圖2E)。純種肉雞(257,16.2%)的ROH數(shù)和平均基因組覆蓋率均高于本地雞(103,7.0%)和GGS(圖2E)。
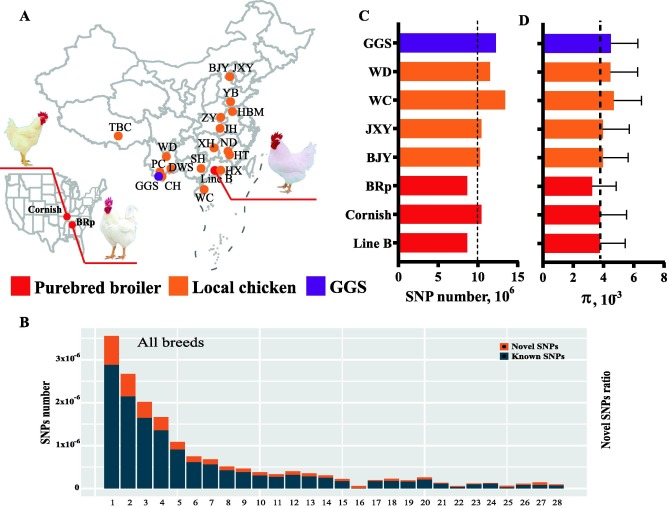
圖2 種群遺傳多樣性與數(shù)量統(tǒng)計(jì)學(xué)歷史推斷
馴化和人工選擇后的遺傳負(fù)荷
馴化會(huì)導(dǎo)致有害突變在雞和其他家畜中積累。表1計(jì)算了每個(gè)基因組中有害的和同義的SNP的數(shù)量和頻率。結(jié)果顯示,在全基因組中,純種肉雞和地方雞的有害SNP(dSNP)分別比GGS多15.63%和24.05%。雖然3組間同義SNP的數(shù)量存在顯著差異,但肉雞和地方雞的dSNP與同義SNP的比例和dSNP的頻率高于GGS。總之,由于馴化和定向育種,在地方雞和肉雞中發(fā)現(xiàn)了快速積累的dSNP。接下來,通過比較雜合子和純合子dSNP的水平,發(fā)現(xiàn)肉雞和地方雞品種中分別有56.75%和69.85%的dSNP保持雜合子狀態(tài),而GGS中的dSNP(50.90%)。與當(dāng)?shù)仉u品種相比,純種肉雞攜帶更多的純合子(肉雞與當(dāng)?shù)仉u544),雜合子較少(肉雞與當(dāng)?shù)仉u1427,1885),純合和雜合dsnp的比例與當(dāng)?shù)仉u品種之間表現(xiàn)出相似的趨勢(表1)。這些結(jié)果表明,dSNP主要是在雜合狀態(tài)下積累的,部分有害的SNP由于集約化養(yǎng)殖而消除。純種肉雞的基因組特征
為了推斷現(xiàn)代肉雞選擇性掃描的基因組位點(diǎn),使用了基于XP-CLR和ZLSBL測試的等位基因頻率分化。結(jié)合這四種方法定義候選發(fā)散區(qū)域(四種方法中至少需要兩個(gè)信號(hào)),窗口大小為40 kb,步長為20 kb(圖3A-B)。通過結(jié)合不同方法的結(jié)果,在純種肉雞群體中鑒定出包含163個(gè)蛋白質(zhì)編碼基因和58個(gè)lncRNAs的391個(gè)窗口列表(圖3A-B),58個(gè)lncRNAs被注釋為新基因。將育種相關(guān)的選擇性掃描與已知的QTL進(jìn)行比較,發(fā)現(xiàn)選擇的ZLSBL分?jǐn)?shù)和XP-CLR值較高但多樣性降低的區(qū)域涉及與肉類生產(chǎn)、免疫和肉質(zhì)相關(guān)的QTL(圖3C),反映了人類對肉類生產(chǎn)的需求以及育種方案下雞抗病能力的變化。在肉雞中,在GGA18上觀察到一個(gè)導(dǎo)聯(lián)信號(hào),該信號(hào)注釋了肌球蛋白重鏈(MYH)基因家族成員(圖3A-B, D)。MYH1A、MYH1B、MYH1C等是肌球蛋白的組成成分和快肌纖維的標(biāo)記物,參與骨骼肌的發(fā)育過程。此外,發(fā)現(xiàn)3個(gè)SNP在純種肉雞、地方雞和GGS之間存在明顯差異(圖3E)。例如,在純種肉雞中固定SNP rs740451868突變等位基因,而在其他動(dòng)物中固定野生等位基因(圖3F)。除MYH基因家族外,肌肉發(fā)育相關(guān)基因IGF-1、INSR和SOX6被檢測到包含前1%的顯著分化區(qū)域。還發(fā)現(xiàn)了一份包含頂級選擇性掃窗的基因列表,這些基因在功能上對肉雞的馴化和定向育種是合理的。例如,TACR3和SLC26A8參與精子發(fā)生和運(yùn)動(dòng);CPAMD8和INHBA與眼睛發(fā)育有關(guān);GLI3參與了雞的色素沉著。基于表型或生理過程,GO術(shù)語被分為幾類,包括心臟發(fā)育過程(如GO:0008016、GO:0060047和GO:0060421)、肌肉功能(如GO:0051155、GO:0014834和GO:0006941)、生殖(如GO:0022412、GO:0046545、GO:0007292)、骨骼發(fā)育(GO:0060348)和感官(GO:1904058、GO:0021618)。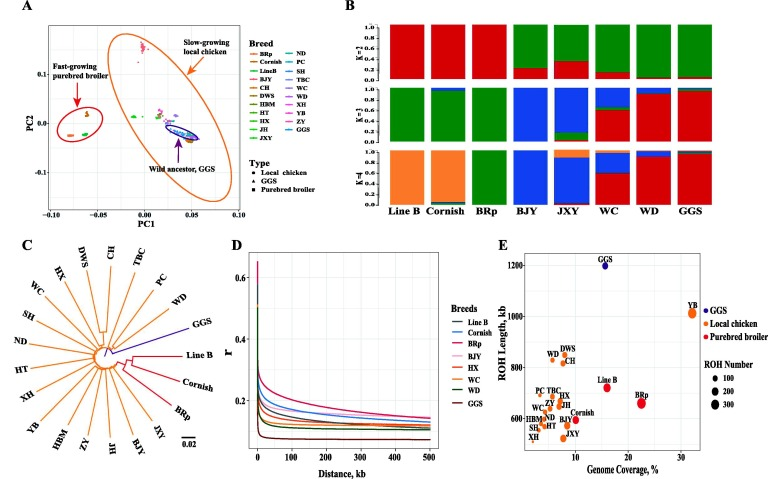
圖3 純種肉雞選擇性掃窗的檢測
純種肉雞和地方雞多組織多階段假定選擇基因(PSGs)的轉(zhuǎn)錄圖譜
為了進(jìn)一步評估肉雞B品種和BJY雞組織中PSGs的表達(dá)豐度,研究者對6種組織進(jìn)行了RNA測序,并計(jì)算了每個(gè)檢測到的基因的計(jì)數(shù)值(圖4A)。基于這兩個(gè)品種的整個(gè)轉(zhuǎn)錄組圖譜,提出了組織特異性的簇。具體來說,具有相似生理功能的組織更容易聚集在一起。在6個(gè)組織中共鑒定出163個(gè)PSG中的83個(gè)DEG(圖4B)。而其他基因似乎在其他組織或發(fā)育階段發(fā)揮作用。通過富集分析,作者發(fā)現(xiàn)肉雞B品種和BJY雞之間的緊密連接通路和肌凝蛋白功能有顯著差異。這些功能和基因在B品種中明顯占優(yōu)勢。參與這些功能的基因(MYH1A、MYH1B和MYH1C)主要與肌肉發(fā)育有關(guān),尤其是胸肌。此外,作者還檢測到大多數(shù)假定選擇的lncRNA不表達(dá),只有9個(gè)lncRNA在6個(gè)組織中表達(dá),但沒有發(fā)現(xiàn)顯著差異。
接著,研究者對163個(gè)PSG中的組織特異性基因(TSG)進(jìn)行分析,在兩個(gè)階段分別發(fā)現(xiàn)了18個(gè)和22個(gè)基因(圖4C)。對于這些TSG,超過50%的基因在肌肉組織中高表達(dá),包括TMEM38A和GGA18上的MYH基因家族成員。此外,TPM4、FTCD、DRAM1、TMEM26也分別在心、肝、肺中特異性表達(dá)。與組織特異性表達(dá)譜一致的是,與BJY雞相比,MYH基因家族中的一些基因(MYH1B、MYH1C、MYH1G)在D1和D42中在B品種的胸肌中高表達(dá)(圖4C)。FTCD和BTBD11在肝臟中表達(dá)水平較低,并在肉雞中特異性表達(dá)(圖4C)。此外,發(fā)現(xiàn)COL6A2在腹部脂肪中高表達(dá),但在肝臟中有差異表達(dá)。此外,作者還發(fā)現(xiàn)SOX6基因在B品種胸肌中的表達(dá)高于BJY雞。綜上所述,B品種各器官與BJY雞有明顯差異,可以推斷,在轉(zhuǎn)錄水平上,肉雞與本地雞種的肌肉存在主要差異。MYH基因家族中的基因(MYH1A、MYH1B、MYH1C等)被確定為調(diào)節(jié)肌肉發(fā)育的功能靶點(diǎn)。隨后,研究者通過加權(quán)基因共表達(dá)分析網(wǎng)絡(luò)(WGCNA)篩選出15620個(gè)基因。根據(jù)整個(gè)轉(zhuǎn)錄組譜,共收集到20個(gè)模塊(圖4D-E),除灰色模塊外,每個(gè)模塊包含64-5513個(gè)基因。為了獲得組織特異性模塊,以相關(guān)系數(shù)r>0.5或r<-0.5,P<1.0-10為標(biāo)準(zhǔn),評價(jià)20個(gè)模塊與6個(gè)組織的相關(guān)性。其中鑒定出10個(gè)組織特異性模塊(圖4E)。例如,暗藍(lán)綠色和綠黃色模塊與胸肌高度相關(guān)。根據(jù)GO注釋,研究者發(fā)現(xiàn)基因在組織特異性模塊中的功能富集與組織功能一致。例如,胸肌特異性模塊中的基因在調(diào)節(jié)成肌細(xì)胞分化和骨骼肌細(xì)胞分化以及糖酵解過程和糖異生中富集(圖4F)。白色和黃色模塊對應(yīng)肝臟,這些模塊中的基因與氧化還原酶活性、氧化還原過程以及脂肪酸代謝有關(guān)。此外,根據(jù)GS>0.2和MM>0.8的標(biāo)準(zhǔn),近23%參與組織特異性模塊的基因被確定為與這6個(gè)組織相關(guān)的核心基因。例如,核心基因的一個(gè)子集,包括MYH1A、MYH1C和MYH1D,被發(fā)現(xiàn)在胸肌中具有高相關(guān)性和特異性表達(dá),這強(qiáng)調(diào)了MYH基因家族成員在胸肌中的作用。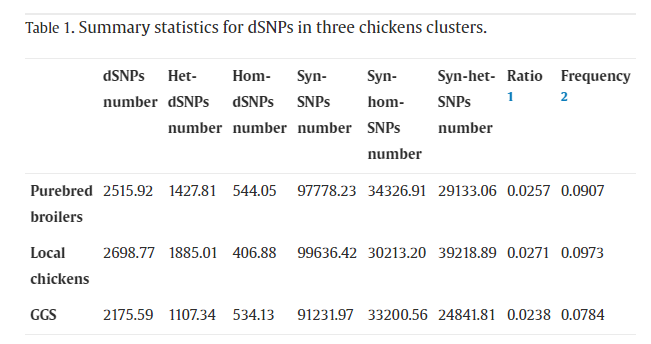
圖4 純種肉雞B系和地方品種BJY雞推定選擇基因(PSGs)轉(zhuǎn)錄組圖譜
GWAS與BrW和BrP的精細(xì)映射
BrW和BrP都是雞育種的關(guān)鍵性狀。純種肉雞B品種經(jīng)過數(shù)代基因組選擇,兩種性狀也發(fā)生了顯著變化。因此,研究者使用GWAS方法在B品種中檢測與它們相關(guān)的遺傳標(biāo)記,結(jié)果發(fā)現(xiàn)位于GGA5上的一個(gè)580 kb的區(qū)域(10.81-11.39 Mb)顯示出兩個(gè)胸肌性狀的顯著峰值(圖5A-B)。根據(jù)Q-Q圖中基因組膨脹因子(0.989 ~ 0.992)的計(jì)算結(jié)果,未發(fā)現(xiàn)群體分層現(xiàn)象。BrW和BrP分別有18個(gè)和36個(gè)SNPs超過提示閾值。對于兩個(gè)性狀GWAS結(jié)果中領(lǐng)先的SNP,基因型遺傳變異占比分別為13.61%和9.57%。在該區(qū)域內(nèi),還發(fā)現(xiàn)了INSC、SOX6和4個(gè)非編碼RNA。隨后,研究者根據(jù)B品種與地方雞種和GGS的比較,進(jìn)行選擇性分析,縮小候選區(qū)域(圖5C),結(jié)果發(fā)現(xiàn)了一個(gè)30kb的區(qū)域有明顯的選擇性特征。為了驗(yàn)證這一結(jié)果,作者還對3只快速生長純種肉雞、本地雞和GGS進(jìn)行了相同的分析。在快速生長的純種肉雞中,存在著相似的選擇性特征,即富集雜合度降低,核苷酸多態(tài)性降低,群體分化更高,而在本地雞和GGS并非如此。因此,縮小的30 kb區(qū)域是一個(gè)有效的候選發(fā)散區(qū),影響了胸肌性狀。研究者在該區(qū)域內(nèi)共檢測到133個(gè)SNP,其中17個(gè)SNP與BrW或BrP相關(guān),并確定了三個(gè)主要的單倍型(圖5D)。攜帶單倍型II的雞胸肌的表型更為突出。單倍型I和單倍型III之間只有一個(gè)堿基不同(圖5D),單倍型I和單倍型III之間沒有表型差異。這種SNP可能不是一個(gè)因果變異,沒有進(jìn)一步考慮。因此,作者分析并確定了兩種主要的單倍型(rHap I和rHap II)(圖5D)。同樣,具有不同單倍型的雞之間也存在顯著的表型差異(圖5E-F)。此外,還確定了其他20個(gè)品種的單倍型狀態(tài)(圖5G);僅檢測到速生純種肉雞攜帶單倍型II,其余18個(gè)肉雞品種均未發(fā)現(xiàn)該單倍型(圖5G)。所有證據(jù)表明,單倍型II可能是肌胸肉性狀的遺傳因素。
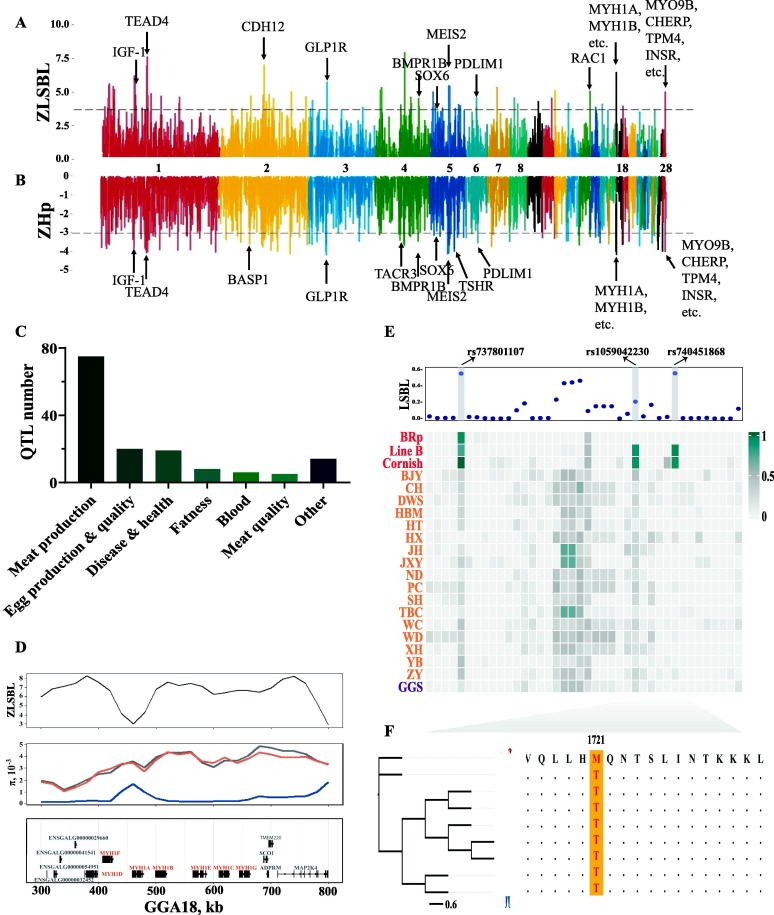
圖5 純種肉雞B系BrW和BrP的GWAS和精細(xì)定位
SOX6基因是BrW和BrP的潛在致病基因
為了確定有效基因,研究者首先對胸肌中的候選基因進(jìn)行了RT-PCR檢測。從比較結(jié)果來看,根據(jù)不同的單倍型,只有SOX6在mRNA和蛋白水平上有差異表達(dá)(圖6A)。雖然INSC基因沒有差異表達(dá),但這四種lncRNA在胸肌肉樣本中沒有表達(dá)。為了最大化挖掘致病基因的可能性,作者檢測了候選區(qū)域周圍基因的表達(dá)。然而,在兩個(gè)精制單倍型樣品之間沒有發(fā)現(xiàn)DEG。此外,作者還確定了不同發(fā)育階段的表達(dá)變化,發(fā)現(xiàn)SOX6隨生長發(fā)育過程呈上升趨勢,其他基因表達(dá)波動(dòng)較小(圖6B)。因此,SOX6被鑒定為可能影響BrW和BrP的功能基因。為了進(jìn)一步驗(yàn)證,研究者首先進(jìn)行了基于SOX6周圍SNP的cis-eQTL分析。結(jié)果發(fā)現(xiàn)盡管它們并不有助于縮小區(qū)域或單倍型,但是對SOX6表達(dá)有顯著影響(圖6C)。接下來,作者通過SMR來估計(jì)SOX6對胸肌性狀的影響。SMR共發(fā)現(xiàn)14個(gè)合格的SNPs,敏感性分析證實(shí)不存在異質(zhì)性(圖6D),且SOX6和胸肌性狀之間的關(guān)聯(lián)不受任何單個(gè)SNP的驅(qū)動(dòng)。然后,SOX6對BrW和BrP均有顯著的正向作用(圖6E),證明了SOX6是影響胸肌性狀的功能基因。雖然已經(jīng)證實(shí)SOX6的表達(dá)水平與B品種的BrW和BrP表型之間存在一定的關(guān)聯(lián),但還需要在其他品種中驗(yàn)證這種關(guān)聯(lián)。研究者比較了當(dāng)?shù)仄贩NJXY雞、JL雞和純種肉雞B品種的基因表達(dá)和表型變化(圖6F-G)。和預(yù)期一致,SOX6表達(dá)量越低,胸肌尺寸越小,且SOX6表達(dá)量與3個(gè)品種BrW和BrP呈正相關(guān)(圖6G-I)。此外,B品種中SOX6的蛋白水平持續(xù)升高,這與mRNA的測量結(jié)果一致。GWAS表明SOX6可能是調(diào)控木質(zhì)化雞胸肉(WS)和白條紋雞胸肉(WB)的潛在致病基因。因此,研究者對WB和WS進(jìn)行評分,并將其與SOX6相關(guān)聯(lián),以研究其調(diào)控模式。在B品種中,SOX6與WB和WS呈正相關(guān)(圖6J-K)。CF和肌病之間的關(guān)系已在之前的報(bào)告和本研究中得到證實(shí),并發(fā)現(xiàn)SOX6對CF性狀有加速作用。此外,胸肌損失也與SOX6相關(guān)。因此,研究者發(fā)現(xiàn)SOX6對胸肌病和肉質(zhì)相關(guān)性狀具有至關(guān)重要的影響。
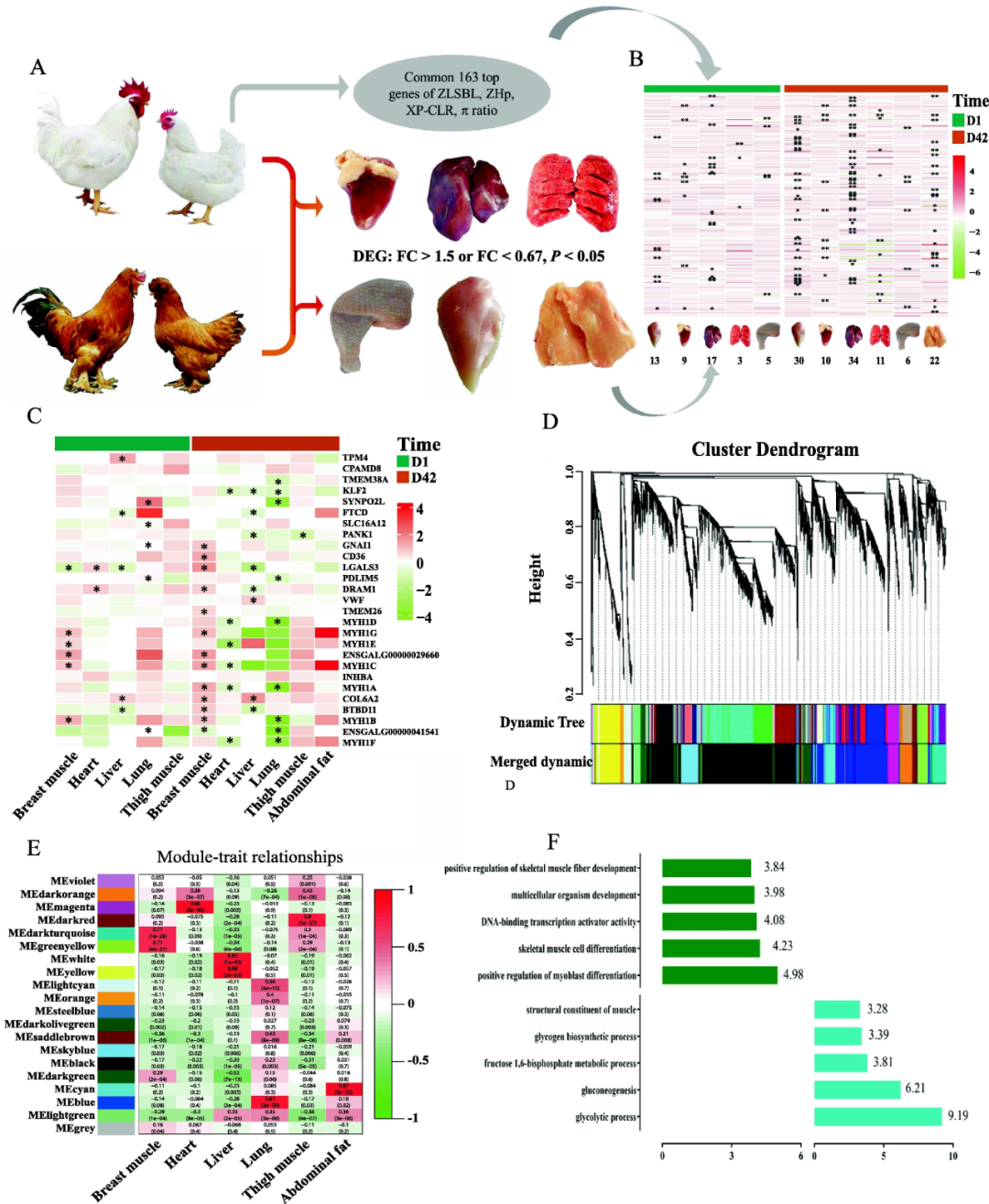
圖6 SOX6基因在不同類型品種中的表達(dá)及表型相關(guān)性分析
【小結(jié)】
本研究構(gòu)建了一個(gè)包含與肌肉發(fā)育相關(guān)的多個(gè)組織和階段的基因組變異和轉(zhuǎn)錄組變異的大規(guī)模圖譜,為探索肉雞生產(chǎn)性狀的遺傳基礎(chǔ)提供了充分和有價(jià)值的信息。根據(jù)基因組和轉(zhuǎn)錄組學(xué)證據(jù),胸肌是純種肉雞與當(dāng)?shù)仉u或GGS的主要差異。PSGs、TSGs和DEGs有助于更好地理解不同類型雞的進(jìn)化差異。SOX6-MYH1s和BrW、BrP及肌病的致病基因SOX6將有助于針對雞的高肉產(chǎn)量進(jìn)行選擇。