










熱脅迫是植物最具挑戰性的環境因子之一,植物已經進化出各種機制來應對它。然而,熱應激引起的脂質重塑的作用尚未完全了解。本研究結合基于UPLC-QTOF/ MS的脂質組學和轉錄組學分析,揭示了熱脅迫下蓖麻脂質重塑的分子基礎。作者在蓖麻幼苗中檢測到297種脂類化合物,其中54種在高溫脅迫下表現出豐度變化。特別是多不飽和三酰基甘油(TAG)和甘油二酰基(DAG)顯著增加,而多不飽和單乳糖二酰基甘油(MGDG)顯著減少。當熱應激減輕后,這些脂質分子恢復到正常水平。作者發現熱誘導的標簽主要在細胞質中積累。轉錄組學分析表明,用于標記的多不飽和脂肪酸(FAs)并非來自從頭合成,而是可能來自脂質重塑通過脂肪酶活性,并隨后通過二酰基甘油酰基轉移酶(DGAT)酯化成標記。該研究的結果表明,TAG可能作為中間體脂質轉換提供了植物應對熱脅迫的一種機制,拓寬了對植物熱脅迫適應過程中脂質重塑的理解。
該論文“Integrated lipidomic and transcriptomic analysis reveals triacylglycerol accumulation in castor bean seedlings under heat stress ”于2022年2月25日,由西北大學與中科院昆明植物所的研究人員合作在Industrial Crops & Products發表。


研究材料
蓖麻變種ZB306種子表面消毒后,置于濕濾紙上,25℃萌發。2天后,將萌發的種子移栽到盆栽土壤中,光照16小時/8小時循環,25℃/22℃,在溫室中生長。將4周大的幼苗分為兩組,每組20株。將25℃光照12 h的幼苗定義為對照組(以下簡稱CK),將45℃光照12 h的幼苗定義為熱應激組(簡稱HS)。熱處理后,半數幼苗恢復正常(25℃12 h-night),定義為恢復組(RHS)。每組幼苗分別采集,立即用液氮冷凍保存在-80℃的超低溫冰箱中進行后續分析。
技術方法
脂質薄層色譜分析,LC-MS質譜分析,亞細胞定位,轉錄組測序,qRT-PCR。
研究結果

與CK組的幼苗相比,HS幼苗沒有表現出明顯的表型差異,表明蓖麻幼苗對熱脅迫具有很強的耐受性。作者采用脂質薄層色譜分析發現熱應激可以誘導蓖麻幼苗中TAG的積累和MGDG的減少。
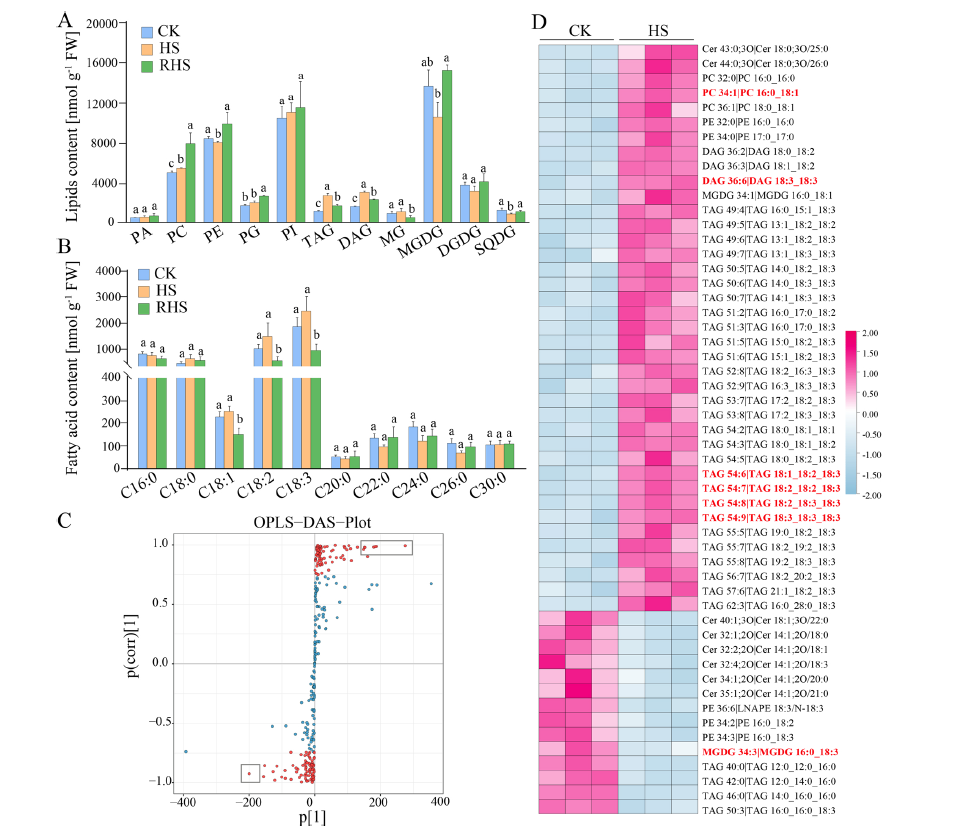
多重比較表明,與CK組相比熱處理導致 TAG、DAG 和 PC 含量顯著增加,MGDG、SQDG 和 PE 含量顯著降低,而PA,PG和PI含量在熱處理后有略微增加,在熱應激解除后,SQDG和PE的含量恢復到與CK相當的水平,TAG和DAG相對于HS顯著降低,但仍高于CK幼苗。在從熱處理中恢復后,PC 的水平持續增加,而 RHS 中的 MGDG 含量恢復到高于 HS 或 CK 的水平。此外,作者觀察到熱應激并沒有引起總FFA含量的明顯變化,但不飽和FAs的含量顯著降低。此外,對不同脂質分子的鑒定顯示,有39種脂質表達上調,15種脂質表達下調,值得注意的是,在熱應激下,DAG和TAG顯著積累,增加了2- 43倍,尤其是DAG6:6、TAG54:6、TAG54:7、TAG54:8和TAG54:9。
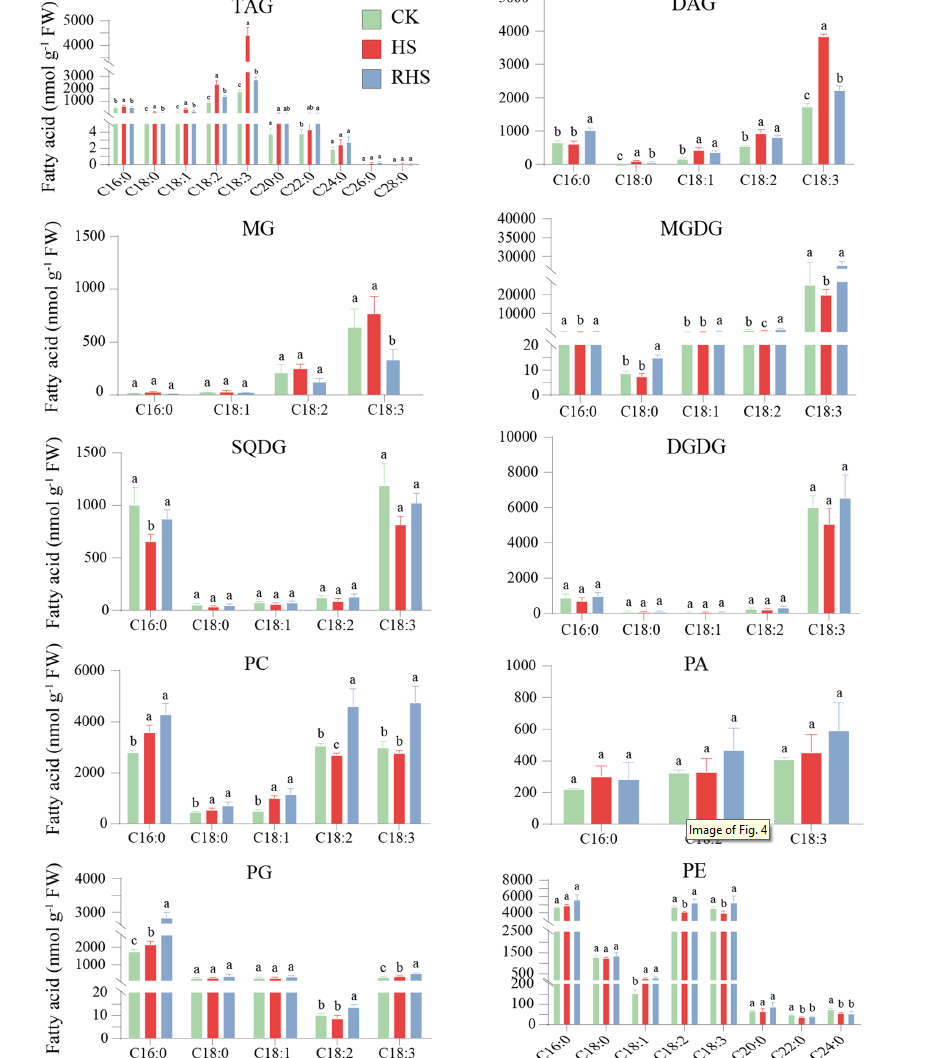
作者分析了與上述不同脂質化合物酯化的 FA 物種。發現 PA、PC、PE、TAG 和 DAG 含有相似的 FA 種類,主要由 C16:0、C18:2 和 C18:3 組成,而 PG 主要由 C16:0 組成,約占總數的 68% FA。MGDG 和 DGDG 主要由 C18:3 組成,分別占總 FA 的 93% 和 82%。另一方面,MG主要含有C18:3(~72%),SQDG主要由C16:0(~41%)和C1:3(~49%)組成。
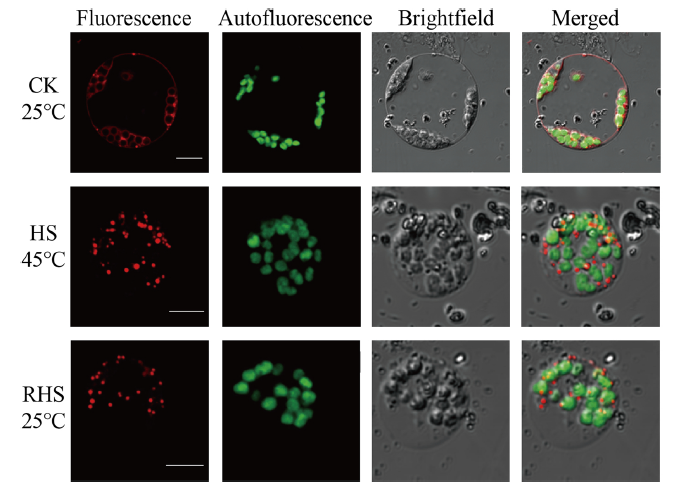
為了研究熱誘導的TAGs、具體積累的部位,作者用一種叫尼羅紅的親脂質染料進行熒光定位,發現在熱處理后的葉片獲得的原生質體中產生了大量的脂滴(紅色熒光),而在從CK獲得的原生質體中僅觀察到少量脂滴。熱應激恢復后,脂滴數量相對于HS明顯減少,但仍高于CK(見圖5),與TAG含量的變化一致。
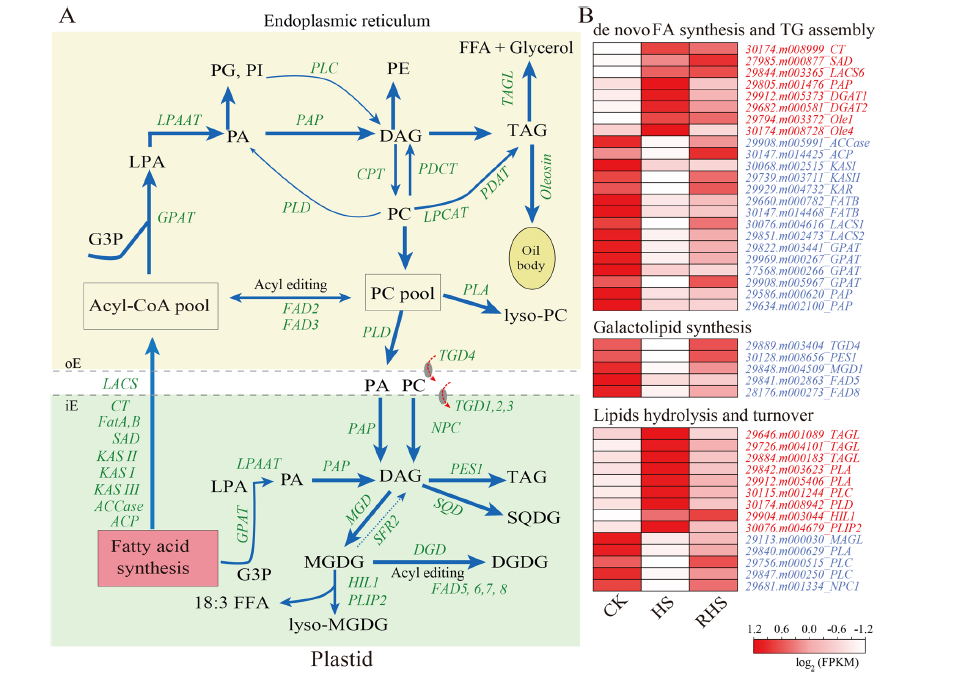
作者分析了熱脅迫下TAG積累的分子學基礎,通過檢測蓖麻基因組作者確定了 135 個基因,這些基因編碼參與 FA 合成等。在熱脅迫下蓖麻幼苗中17個基因表達上調,25個基因表達下調。其中參與FA合成的關鍵基因,如ACCase、3- keto酰基acp合成酶I (KAS I)、3- keto酰基acp還原酶(KAR)、棕櫚酰酰基載體蛋白硫酯酶(FATB)和LACS1在熱應激下顯著下調。
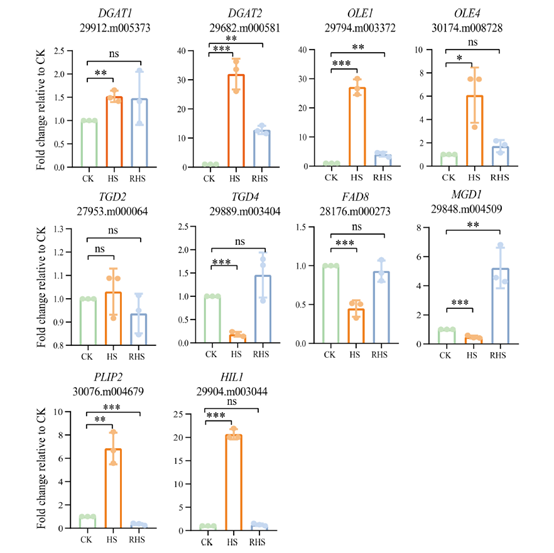
作者還選擇了6個上調基因(DGAT1、DGAT2、OLE1、OLE4、PLIP2和HIL1)和4個下調基因(TGD2、TGD4、FAD8和MGD1)進行qRT-PCR驗證。所有上調的基因以及4個下調的基因中有3個得到了qRT-PCR的證實,TGD2在熱脅迫下的表達沒有發生變化。這些結果表明,熱誘導的轉錄水平的變化協調了脂質代謝的變化。
作者對熱應激和恢復過程中蓖麻幼苗的脂質組學和轉錄組學進行了綜合分析,并鑒定了大量熱響應脂質分子和基因。發現熱應激可顯著誘導 TAGs 合成并儲存在胞質溶膠中,但在恢復過程中它們會減少。TAG 可能是回收從其他脂質釋放的 FA 的瞬時中間體,尤其是葉綠體脂質,如 MGDG。轉錄組分析確定了許多與協調脂質重塑的熱反應性脂質相關的基因。該研究結果增加了對熱應激下蓖麻和其他植物物種脂質周轉的理解,同時為操縱特定脂質分子,尤其是 TAG 提供了潛在的應用,以提高植物的耐熱性。

